Caractéristiques caractéristiques des espèces r et k. Stratégies végétales écologiques
Comment déterminer la valeur d’un individu pour une population ?
« La sélection naturelle ne reconnaît qu'un seul type de « monnaie » : une progéniture prospère" (E. Pianka, 1981).
Nous avons dit qu'une population est une entité potentiellement immortelle composée d'individus mortels. Pour maintenir l’existence d’une population, un individu doit survivre à lui-même et laisser une descendance qui peut également survivre. Notez la dualité de cette tâche. La plus grande chance de survie sera probablement celle qui ne dépensera pas ses ressources et l'énergie qu'elle en tirera pour produire une progéniture. Mais un peu de temps passera et un tel individu disparaîtra de la population sans laisser de trace. Au « pôle » opposé se trouve un individu hypothétique qui, immédiatement après son apparition, commence à consacrer toute son énergie à la production de descendants. Une telle créature mourra elle-même et, si ses descendants héritent d’une manière tout aussi inefficace d’allocation des ressources, elle produira des descendants qui n’auront aucune chance de survie.
Cela signifie que l’individu qui combine les coûts de sa propre survie et la production d’une progéniture dans une combinaison optimale devrait avoir la plus grande valeur pour la population. Il est possible d’évaluer à quel point cette combinaison est optimale. Pour ce faire, il faut calculer sous quelle combinaison, dans des conditions données, un individu laissera la plus grande contribution possible à la génération future. La mesure utilisée à cet effet en biologie mathématique des populations s'appelle valeur reproductive. La valeur reproductive est une mesure généralisée de la survie et de la fertilité qui prend en compte la contribution relative d'un organisme aux générations futures.
« Il est facile de décrire un organisme hypothétique possédant toutes les caractéristiques nécessaires pour atteindre une valeur reproductive élevée. Il se reproduit presque immédiatement après la naissance, produit une progéniture nombreuse, de grande taille et protégée, dont il prend soin ; il se reproduit de nombreuses fois et souvent au cours d'une longue vie ; il remporte la compétition, évite les prédateurs et obtient facilement de la nourriture. Il est facile de décrire une telle créature, mais difficile de l'imaginer...." (Bigon et al., 1989).
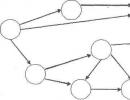
Vous comprenez qu'une telle impossibilité naît de l'incohérence des tâches d'auto-entretien et de reproduction (Fig. 4.15.1). L'un des premiers à s'en rendre compte fut en 1870 le philosophe anglais Herbert Spencer, qui parlait de l'alternative selon laquelle le corps maintiendrait sa propre existence et se perpétuerait dans ses descendants. En langage moderne, on peut dire que ces paramètres sont liés par des corrélations négatives, relation dans laquelle l'amélioration du système dans un paramètre doit s'accompagner de sa détérioration dans un autre.
Riz. 4.15.1. Chez le rotifère Asplanchna les chances de survie diminuent à mesure que la fécondité augmente (Pianka, 1981)
Différentes espèces (et différentes populations) répartissent l'énergie différemment entre l'auto-entretien et la reproduction. On peut parler d'une stratégie d'espèce, exprimée dans la manière dont les représentants de l'espèce obtiennent des ressources et comment ils les dépensent. Seule une stratégie dans laquelle les individus reçoivent une quantité suffisante d'énergie pour pouvoir grandir, se reproduire et compenser toutes les pertes dues à l'activité des prédateurs et à divers malheurs peut réussir.
Les traits liés à différentes stratégies adaptatives peuvent être liés par la relation compromis, c'est-à-dire des corrélations négatives irrésistibles (relation soit-soit). Ainsi, le ratio de compromis relie le nombre de descendants et leur taux de survie, leur taux de croissance et leur résistance au stress, etc. Les écologistes américains R. MacArthur et E. Wilson ont décrit en 1967 deux types de stratégies d'espèces, qui sont le résultat de deux types différents de sélection et sont liées par une relation de compromis. Les notations acceptées pour ces stratégies (r- et K-) sont tirées de l'équation logistique.
Selon le modèle logistique, deux phases peuvent être distinguées dans la croissance démographique : avec une croissance accélérée et avec une croissance décélérée (Fig. 4.15.2). Au revoir N est faible, la croissance démographique est principalement influencée par le facteur rN et la croissance démographique s’accélère. A cette phase ( phase r) la croissance démographique s'accélère, et son nombre est d'autant plus élevé que la capacité des individus à se reproduire est élevée. Quand N devient assez élevée, la taille de la population commence à être principalement influencée par le facteur (K-N)/K. A cette phase ( Phase K) la croissance démographique ralentit. Quand N=K, (K-N)/K=0 et la croissance démographique s’arrête. À la phase K, plus le paramètre est élevé, plus la taille de la population est élevée K. Plus les individus sont compétitifs, plus il est élevé.

Riz. 4.15.2. Phases r et K de la croissance démographique conformément au modèle logistique
On peut supposer que les populations de certaines espèces sont la plupart du temps en phase R. Chez ces espèces, la valeur reproductive maximale est donnée aux individus capables de se reproduire rapidement et de capturer un environnement vide avec leurs descendants. En d'autres termes, à cette phase, la sélection contribuera à une augmentation du paramètre r- le potentiel reproducteur. Cette sélection s'appelle r-sélection, et les espèces résultantes - g-stratèges.
Pour les espèces dont les populations sont la plupart du temps en phase K, la situation est complètement différente. La valeur reproductive maximale de ces populations sera inhérente aux individus qui seront si compétitifs qu'ils pourront obtenir leur part de la ressource même dans des conditions de rareté ; ce n’est qu’alors qu’ils pourront se reproduire et contribuer à la génération suivante. Une population composée de tels individus aura plus haute valeur paramètre K- la capacité de l'environnement que celle composée d'individus qui ne « savent pas » lutter pour les ressources manquantes. A ce stade, la sélection K agit sur la population, ce qui se traduit par l'émergence d'espèces - K-stratèges. Sélection K vise à augmenter les coûts de développement de chaque individu et à accroître sa compétitivité.
Des transitions entre ces stratégies sont possibles, mais elles sont de nature intermédiaire et ne combinent pas les expressions typiques des deux formes.
« On ne peut pas être à la fois une laitue et un cactus."(E. Pianka).
La dynamique des changements dans la quantité de ressources disponibles et l'intensité de la compétition pour celles-ci sont importantes pour déterminer quelle sélection (r- ou K-) agira sur une espèce. En cas de forte réduction aveugle de la taille de la population causée par un manque de ressources dû à des raisons externes, les stratèges R obtiennent un avantage, et en compétition pour la ressource manquante, les stratèges K obtiennent un avantage.
Le choix entre la stratégie R (augmentation de la fertilité) et la stratégie K (augmentation de la compétitivité) semble assez simple, mais il affecte de nombreux paramètres des organismes et de leurs cycles de vie. Comparons ces stratégies sous leur forme typique (tableau 4.15.1).
Tableau 4.15.1. Caractéristiques de la sélection r et K et des stratégies
|
Caractéristiques |
r-sélection et r-stratèges |
K-sélection et K-stratèges |
|
Modifiable, imprévisible |
Constant, prévisible |
|
|
Mortalité |
Catastrophique, indépendamment de la densité de population |
Causé par la concurrence, dépendant de la densité de population |
|
Courbe de mortalité |
Généralement de type III |
Généralement de type I ou II |
|
Taille de la population |
Modifiable, déséquilibré |
Constante, proche de la capacité maximale du milieu |
|
Ressources gratuites |
L’émergence de ressources gratuites, comblant le « vide écologique » |
Il n'y a presque pas de ressources gratuites, elles sont occupées par des concurrents |
|
Compétition intra- et interspécifique |
||
|
Taille du corps |
Relativement petit |
Relativement large |
|
Développement |
Lent |
|
|
Maturité sexuelle |
||
|
Taux de reproduction |
||
|
Reproduction au cours de la vie |
Souvent ponctuel |
Répété |
|
Descendants dans une couvée |
Peu, souvent seul |
|
|
Montant de la ressource par enfant |
||
|
Durée de vie |
Court |
|
|
Adaptations |
Primitif |
Parfait |
|
Optimisé |
Productivité |
Efficacité |
Il peut être surprenant de savoir pourquoi les stratèges R se caractérisent par une reproduction unique, alors que les stratèges K se caractérisent par une reproduction répétée. Cette fonctionnalité est plus facile à expliquer avec un exemple. Imaginez des souris infestant une grange céréalière (beaucoup de ressources, pas de concurrence). Considérons deux types de stratégies.
Vue n°1. La maturité sexuelle est de 3 mois, le nombre de petits dans la couvée est de 10, la femelle vit un an et est capable de se reproduire tous les trois mois.
Vue n°2. La maturité sexuelle est de 3 mois, le nombre de petits dans la couvée est de 15, après les avoir nourris, la femelle meurt d'épuisement.
Dans le premier cas, après trois mois, 10 petits et leurs parents commenceront à se reproduire (12 animaux au total), et dans le second, jusqu'à 15 petits. Le deuxième type peut fournir un taux plus élevé de capture de ressources gratuites. Une stratégie R typique oblige les individus à se reproduire le plus tôt et le plus durement possible, et les stratèges R sont donc souvent limités à une seule saison de reproduction.
D’un autre côté, il est facile de comprendre pourquoi les stratèges K typiques se reproduisent plusieurs fois. Dans un environnement compétitif, seul le descendant pour le développement duquel de nombreuses ressources ont été dépensées survivra. En revanche, pour survivre et se reproduire, un adulte doit consacrer une quantité d’énergie importante à son propre entretien et à son développement. Par conséquent, dans le cas limite, les stratèges K produisent une progéniture à la fois (comme, par exemple, les éléphants et les baleines et, dans la plupart des cas, les humains). Mais peu importe à quel point ces animaux sont parfaits, deux parents mourront avec le temps. Pour que la population ne s'éteigne pas, un couple de parents doit laisser un couple de descendants survivants et, par conséquent, doit en donner naissance à plus de deux. Si tel est le cas, une condition nécessaire à la survie des stratèges K est la reproduction multiple de leurs individus constitutifs.
En 1935, le botaniste soviétique L.G. Ramensky a identifié trois groupes de plantes, qu'il a appelés coénotypes (le concept de stratégies n'était pas encore formé) : les violentes, les brevets et les explérentes. En 1979, ces mêmes groupes (sous des noms différents) ont été redécouverts par l'écologiste anglais J. Grime (Fig. 4.15.3). Ces stratégies sont les suivantes.

Riz. 4.15.3. « Grime's Triangle » - classification des stratégies spécifiques
- Tapez C (concurrent, concurrent), violent selon Ramenski ; dépense la plupart l'énergie nécessaire à maintenir la vie des organismes adultes, domine dans les communautés durables. Parmi les plantes, ce type comprend le plus souvent les arbres, les arbustes ou les graminées puissantes (par exemple le chêne, le roseau).
- Les types (tolérant au stress, tolérant au stress); patient selon Ramenski ; grâce à des adaptations spéciales, supporte des conditions défavorables ; utilise des ressources là où presque personne ne lui rivalise pour les obtenir. Il s’agit généralement d’organismes à croissance lente (par exemple sphaignes, lichens).
- Tapez R(de lat. ruderis, ruderal), expérimenter selon Ramenski ; remplace les violents dans les communautés détruites ou utilise des ressources temporairement non réclamées par d’autres espèces. Parmi les plantes, ce sont des annuelles ou des bisannuelles qui produisent de nombreuses graines. Ces graines forment une banque de graines dans le sol ou sont capables de se propager efficacement sur une distance considérable (par exemple pissenlit, épilobe). Cela permet à ces usines d'attendre que les ressources soient libérées ou de reprendre à temps les zones libres.
De nombreuses espèces sont capables de combiner différents types de stratégies. Le pin appartient à la catégorie CS car il pousse bien sur des sols pauvres sols sableux. L'ortie est un stratège en CR car elle domine les habitats perturbés.
La stratégie d'une espèce peut être flexible. Chêne anglais - violent dans la zone forêts de feuillus et patient dans steppe du sud. La technologie japonaise du bonsaï (culture d’arbres nains en pot) peut être présentée comme un moyen de transformer les violents en patients.
Une tâche intéressante consiste à comparer les stratégies selon MacArthur-Wilson et selon Ramensky-Grime. Il est clair que les r-stratèges correspondent aux organismes de type R, les explérents. Mais les stratèges K correspondent non seulement aux organismes de type C, les violents, mais aussi à ceux qui appartiennent au type S, les patients. Les violents maximisent leur compétitivité (et la capacité de l'environnement) dans des conditions de concurrence intense pour les ressources favorables à la consommation, et les patients - dans des conditions de consommation difficile des ressources. En d’autres termes, les problèmes résolus par un chêne rivalisant pour la lumière dans une forêt dense et par une fougère survivant dans une pénombre au fond d’une grotte ont beaucoup en commun : la nécessité d’optimiser la consommation des ressources et d’améliorer la condition physique individuelle.
Histoire développement du concept de « stratégie écologique » chez les plantes .
Premièrement, le terme « stratégie » désignait un ensemble de propriétés qui aident les organismes à survivre dans des conditions données et ne s’appliquait qu’aux organismes animaux.
Les stratégies R et K ont été distinguées sur la base du rapport entre les coûts de reproduction et d'entretien de la progéniture.
Les stratèges K se distinguent par le fait qu'ils s'occupent d'un petit nombre de descendants, comme par exemple chez les éléphants. Les stratèges R se caractérisent par une fertilité maximale et un manque de soins pour la progéniture, par exemple les vers ronds.
Propriétés du K-iR.- les stratégies chez les animaux.
| R.-stratégie | Stratégie K |
| Caractérisé par un développement rapide des individus | Caractérisé par un développement lent des individus |
| Haute fertilité | Faible fertilité |
| Petite taille des individus | Grande taille d'individus |
| Durée de vie courte | Espérance de vie importante |
| Actes de reproduction antérieurs | Reproduction tardive |
| Tous les signes visent une productivité plus élevée | Tous les panneaux visent une utilisation efficace des ressources |
| Caractéristique des changements catastrophiques de l'environnement lorsque des biotopes non comblés sont peuplés. | Plus efficace dans un environnement compétitif. |
Plus tard, le terme « stratégie écologique » a commencé à être utilisé en relation avec les organismes végétaux. (20).
Pour la littérature russe, le terme « stratégie » en relation avec les plantes est assez nouveau et a été utilisé pour la première fois par T.A. Rabotnov (1975), qui a nommé L.G. isolé. Ramensky (1936) « types coénobiotiques ».
Rabotnov a proposé de comprendre la stratégie d'une espèce comme « un ensemble d'adaptations qui lui donnent la possibilité de vivre avec d'autres organismes et d'occuper une certaine place dans la biogéocénose correspondante ». (10)
McLeod a été le premier à attirer l'attention sur la présence de conditions préalables dans les plantes qui déterminent leur statut dans la communauté, en 1894, qui a divisé toutes les espèces en « capitalistes » et « prolétaires ».
Cependant, tant l'analogie avec la société elle-même que le principal critère de distinction des types - pollinisation croisée et autopollinisation - n'ont pas abouti, bien que le scientifique ait tenté de rendre les évaluations globales et ait écrit que les « capitalistes » se caractérisent par la présence d'une réserve. nutriments, polycarpicité, intolérance à l'ombrage, etc.
Cette question a connu un développement brillant dans les travaux de Ramensky, publiés dans les années 30, où il a écrit sur 3 types de plantes, qu'il a qualifiées de violentes, de brevets et d'explérentes et les a comparées à des lions, des chameaux et des chacals.
40 ans plus tard, la monographie de J. Grime « Plant Strategies and Processes in Vegetation » était publiée en Angleterre. , dans lequel l’auteur, ne connaissant pas les travaux de Ramensky, a redécrit les trois mêmes types de stratégies appelées compétiteurs, tolérants au stress et ruderaux.
Pour comprendre le type de stratégies, beaucoup a également été fait par E. Pianka, R. Whittaker et T.A. Rabotnov. (11)
Systèmes de base des stratégies écologiques-cénotiques .
Le système d'E. Pianka.
Le système Pianka, le plus utilisé en écologie, comprend deux types de stratégies associées à la sélection K et à la sélection r (basées sur le rapport des parts des coûts énergétiques pour le maintien des individus adultes et pour les processus de reproduction).
La sélection K est une sélection dans un environnement constant (prévisible), où la majeure partie de l'énergie de la population est dépensée en compétition, et avec la sélection r, le principal poste de dépense énergétique est la reproduction.
Le système est le résultat du développement d'idées formulées plus tôt par R.Kh. MacArthur et E.O. Wilson, cependant, c'est E. Pianka qui a analysé en profondeur les conséquences qui résultent de la mise en œuvre de deux types de sélection.
Deux types de stratégie de Pianka sont les plus répandus dans le monde végétal. Et même l'émergence d'hétérospores dans les mousses ou les fougères peut finalement être considérée comme un remplacement de la stratégie r des isospores par la stratégie K du gamétophyte femelle, qui garantit une meilleure survie de la progéniture et remplace un grand nombre de petites isospores par un nombre limité de mégaspores, fournissant les conditions nécessaires au développement du prothalle femelle.
Les K-stratèges sont confinés à des conditions environnementales plus ou moins stables, ont des populations en équilibre où la mortalité est régulée par la densité et sont adaptés à des conditions de compétition intense. Ils sont généralement polycarpiques avec un développement lent et une forme de vie allant des herbes aux arbres. Dans les séries de succession, ces espèces augmentent leur participation à mesure que l'étape de succession approche de son apogée.
Les r-stratèges préfèrent au contraire les habitats instables caractérisés par des populations hors équilibre, dont la mortalité ne dépend pas ou peu de la densité. La concurrence entre ces plantes est faible : ce sont de jeunes plantes monocarpiques, généralement des herbes, moins souvent des arbustes. Dans les séries successorales, ils sont associés aux stades pionniers et ne jouent pas de rôle significatif dans les communautés matures précédant le climax.
Ainsi, le système de types d'E. Pianka est simple - unidimensionnel, mais il correspond pleinement à la perception continue des types.
Il note la relativité de la division de tous les types en 2 types de stratégies, soulignant que le monde n'est pas peint uniquement en noir et blanc et que les options extrêmes, en règle générale, sont associées à toute une série de transitions (E. Pianka, 1981, p.138). (13)
Système R. Whittaker.
R. Whittaker (1980) distingue non pas 2, mais trois types de stratégies, désignées lettres K,r et L. Son système est basé sur les modèles de fluctuations des effectifs de population entre deux limites : K-limite supérieure, correspondant à la densité de saturation maximale et L-limite inférieure, signifiant un certain « zéro de population », correspondant à un nombre qui est incapable d'assurer la survie de la population.
Les stratèges K s’efforcent d’atteindre le niveau K, en y parvenant d’abord grâce à une différenciation extrême des niches. La sélection K affecte les mécanismes par lesquels ils maintiennent leur population dans le processus de compétition et d'autres interactions dans les limites de l'environnement qu'ils occupent. La taille de la population diminue considérablement, mais la tendance générale de ces populations est de fluctuer autour du niveau K.
Le deuxième groupe de populations_r-stratèges. Ils se caractérisent par de fortes fluctuations entre les niveaux de K et L. Ces populations sont instables et ne survivent que grâce au taux élevé de production de diaspores, elles sont mal adaptées aux conditions de compétition intense et aux conditions défavorables qui provoquent du stress.
Le troisième groupe de populations est celui des stratèges L, qui fluctuent autour de la limite inférieure des nombres L, bien qu’ils puissent parfois augmenter leur nombre de manière explosive. Dans de telles populations, la sélection tend à améliorer le mécanisme permettant de survivre aux périodes défavorables, et le taux de reproduction peut être élevé ou non.
Tout en distinguant trois types de sélection avec leur résultat - trois types primaires, Whittaker, comme Pianka, n'a pas en même temps rendu son système absolu.
Si l’on compare les systèmes de Whittaker et de Pianki, il est évident que ses types K et r correspondent aux K et r de Pianki et qu’en effet la différenciation de niche se produit sous l’influence de la sélection K. Il s'agit principalement d'espèces pérennes, se reproduisant souvent par voie végétative et dépensant relativement peu d'énergie dans la sphère générative.
Les plantes ruderales, au contraire, sont raccourcies cycle de vie et une productivité élevée des semences, et donc les coûts de reproduction sont ici plus élevés. C'est une conséquence de la r-sélection.
Le groupe L occupe une position transitoire, puisque les annuelles du désert font partie des plantes éphémères avec un cycle de développement très rapide et une productivité de graines élevée (résultat de la sélection r), mais les arbustes, ainsi que certaines plantes herbacées à gazon, subissent un stress à l'état végétatif. et représentent donc le résultat de l'action de K - sélection (10)
Système Ramensky-Grime.
Ramensky a proposé un système de trois types. Il distingue trois « types coénobiotiques ».
Le premier type, qu'il appelle violents ou « lions », se caractérise par la capacité de s'emparer vigoureusement du territoire, l'intégralité des ressources utilisées et une puissante répression compétitive des rivaux.
Le deuxième type - les patients ou « chameaux » se distinguent par leur capacité à résister à des conditions environnementales extrêmes, c'est-à-dire leur endurance.
Le troisième type - les explerents ou les chacals ne se distinguent ni par leur résistance aux situations stressantes ni par leur pouvoir compétitif élevé, mais sont capables de combler rapidement les écarts entre des usines plus fortes, et lorsqu'elles sont fermées, ils sont également facilement expulsés. (13)
Par la suite, la présentation et le classement de L.G. Ramensky (1935-38) ont été développés par T.A. Rabotnov. (1966, 1975, 1978, 1980). Il a montré la nature complexe de la patience (tolérance au stress) chez les plantes et identifié des patients environnementaux et phytocénotiques.
Les premiers sont capables d'exister dans des conditions défavorables dues à une spécialisation écologique (sur substrats salins, acides, secs, rocheux, etc.) et correspondent le plus aux patients L.G. Ramenski. Ils ont les mêmes optimums autecologiques et synécologiques.
Ces derniers sont capables de survivre longtemps sous la pression des violents dans des conditions écologiquement optimales en maximisant la réduction des processus vitaux. Leurs optimums synécologiques et autecologiques ne coïncident généralement pas. (6 )
Nous trouvons un développement plus approfondi des idées sur les types de stratégies dans de nombreux travaux de J. Grime (J. Grime, 1974, 1978, 1979).
Il en propose essentiellement 3, les mêmes que L.G. Ramensky, type de stratégies écologiques-cénotiques, appelant ces types : concurrents, tolérants au stress et rudéraux (K, S et R, respectivement).
Les stratégies de contacts sociaux les plus clairement opposées se manifestent bien entendu dans le comportement reproductif, c'est-à-dire dans la stratégie d'élevage.
Chez la plupart des espèces, y compris les humains, les deux stratégies de reproduction sont retrouvées. La direction générale de l'évolution humaine peut être définie comme un mouvement allant de r-des stratégies pour K- stratégies. Vous pouvez même indiquer l'heure approximative à laquelle K- la stratégie a commencé à prévaloir - nous sommes au 3ème millénaire avant JC, lorsque le mythe du conflit entre Niobe et Latone est né sur le territoire de l'Asie Mineure.
Niobé a refusé de faire des sacrifices à Latone et à ses enfants, de Zeus à Apollon et Artémis. Elle a expliqué cela, en partie, par le fait qu'elle a sept fois plus d'enfants que Latona. Offensée, Latone s'est plainte aux enfants. Apollon et Artémis, qui ont défendu leur mère, ont tué tous les Niobides avec des flèches.
La signification biologique de ce mythe est évidente : mieux vaut avoir peu de descendants, mais plus adaptés à environnement, qui en compétition vaincra des individus plus nombreux, mais moins adaptés. Et les plus grandes capacités d'adaptation de la progéniture sont obtenues, comme déjà noté, premièrement, par une sélection minutieuse d'un partenaire reproducteur et, deuxièmement, par des soins attentifs à la progéniture - ce que chez l'homme on appelle l'éducation et la formation.
Dans l'évolution humaine r-la stratégie est progressivement remplacée À-stratégie.
Avantage évolutif transmis à K- stratèges, c'est-à-dire les femmes qui : 1) ont soigneusement choisi leur partenaire reproductif (conjoint) et 2) ont eu un comportement parental prononcé, c'est-à-dire prodigué des soins attentifs aux enfants, les élevait et leur donnait une éducation.
Femme K-le stratège souhaite que le partenaire reproducteur dépense toutes les ressources obtenues pour subvenir uniquement aux besoins de sa progéniture.
Malgré le fait qu'en général les humains soient une espèce monogame (plus précisément, parmi les humains, il y a plus de représentants K-stratégies), il y a souvent des porteurs de la stratégie reproductive opposée, qui sont assez indifférents à leurs enfants. Ces personnes, en particulier les femmes, ressentent souvent douloureusement leur indifférence, se considérant coupables du manque de sentiments parentaux. Les médecins classent cette condition comme une névrose particulière de « mauvaise mère ».
Le type de stratégie reproductive à laquelle appartient une personne n'est révélé qu'après la naissance de son enfant. Ensuite, la réaction hormonale qui accompagne l’accouchement initie un complexe de comportements parentaux. Il est difficile de déterminer si une femme appartient à l'un ou l'autre type psychologique avant d'accoucher. -r- ou À- stratégies. Vous ne pouvez pas attirer l'attention sur vos propres enfants.
La froideur ou l'hostilité d'une femme envers ses enfants sont des variantes de la norme. C'est une manifestation extrême r-les stratégies de reproduction.
Si une femme en bonne santé a haut niveau cortisol au repos, c'est-à-dire Si elle appartient au type psychologique B, cela sert de base pour prédire un comportement parental intensif. La concentration de cortisol dans le sang pendant la grossesse augmente chez toutes les femmes. Mais son augmentation est plus importante chez les femmes qui ont par la suite montré un comportement maternel plus prononcé.
En plus du cortisol, le biais d’affiliation parentale se reflète dans le rapport estradiol/progestérone. Une augmentation progressive de ce ratio du début à la fin de la grossesse est un marqueur À-stratégies.
Concernant la régulation hormonale du comportement paternel, c'est-à-dire On sait très peu de choses sur le comportement parental des hommes. Il existe des preuves que le comportement parental est plus prononcé chez les hommes ayant de faibles niveaux de testostérone et des niveaux élevés de prolactine. Les hommes qui passent beaucoup de temps avec leurs enfants de moins d'un an ont des taux de cortisol et de prolactine dans le sang plus élevés que ceux qui consacrent peu de temps à une telle communication, mais les différences n'atteignent pas le niveau de signification statistique.
Importance pratiqueétudes de marqueurs biologiques K-les stratégies sont évidentes. Une femme a des exigences différentes, largement contradictoires, envers son partenaire sexuel et reproductif. Si un amant devait posséder nombre maximum avantages, alors le mari devrait avoir un minimum d'inconvénients. Et seulement deux qualités positives : gagner de l’argent et bien traiter les enfants. Par conséquent, le problème du choix d’un conjoint sera grandement facilité lorsque des signes biologiques spécifiques de l’inclination d’une personne à adopter un comportement garantissant K- stratégie d'élevage. Malheureusement, ce problème est encore loin d’être résolu.
Il convient de noter que le comportement caractéristique des deux stratégies de reproduction ne se manifeste pas seulement dans les relations avec les enfants et les conjoints. Les stratégies de comportement reproductif constituent un cas particulier des stratégies de contact social.
Choisissez - moi ou ce chat !
Eh bien, je te choisis. Après tout, je te connais depuis longtemps, mais c’est la première fois que je vois ce chat.
E. Ouspenski
Le personnage d'E. Uspensky est évident K- stratège, car si un choix alternatif s’impose, il préfère une personne qu’il connaît bien. Le propriétaire du type psychologique opposé choisira un étranger, car la communication avec lui promet de nouvelles impressions et est plus intéressante.
r- Et K- les stratégies de sélection sont un cas particulier r- et les stratégies K de contacts sociaux.
r- Et K-les stratégies de contacts sociaux peuvent être considérées comme des types psychologiques. Les animaux de type B réagissent activement par des réactions comportementales et endocriniennes au comportement d'un autre animal. Les rats de type A sont indifférents au comportement de leur voisin. Les différences dans le système ocytocine de ces animaux sont très révélatrices. Chez les animaux de type A, l'activité du système ocytocine est deux fois inférieure à celle des animaux de type B. Ainsi, il existe une correspondance avec les différences dans les mécanismes humoraux et les types de contacts sociaux chez les animaux de lignées génétiquement sélectionnées.
Examinons quelques exemples de l'effet de l'ocytocine sur le comportement humain.
Les volontaires ont reçu de l'ocytocine intranasale, ce qui a accru la confiance entre les gens.
De plus, le stress social précoce provoqué par la séparation maternelle entraîne une altération des taux d’ocytocine chez les adultes. Par exemple, chez des singes rhésus élevés isolément de leur mère, à l'âge de 18, 24 et 36 mois, le nombre de contacts sociaux d'affiliation, y compris la durée de l'allorooming, a été considérablement réduit, et le nombre de contacts agonistiques et la durée des actes moteurs stéréotypés a été augmentée. Dans de tels isolats, la concentration d'ocytocine dans le liquide céphalo-rachidien est nettement inférieure à celle des isolats normaux, c'est-à-dire des singes élevés avec leur mère.
Des résultats similaires ont été obtenus dans une étude portant sur des personnes manquant de contact avec leurs parents. Les enfants qui ont été privés de soins maternels depuis leur naissance, lorsqu'ils deviennent adultes, souffrent de troubles émotionnels et de troubles de l'affichage. comportement social. Ils montrent également une activité réduite des systèmes ocytocine et vasopressine 147 . Des perturbations du système ocytocine ont également été constatées chez des enfants privés de la présence de leur père. On sait que les enfants de mères célibataires présentent un risque accru de troubles émotionnels. Chez les hommes adultes ayant grandi sans père, l'effet inhibiteur de l'ocytocine administrée par voie intranasale sur l'augmentation du cortisol dans le sang induite par le stress est affaibli.
Pour résumer la discussion sur la question des stratégies de contacts sociaux humains, il faut dire qu'il existe sans aucun doute deux de ces stratégies : r- Et À-. Ils se manifestent principalement dans les relations avec les enfants, mais aussi dans tous les autres contacts sociaux. K-la stratégie est associée à une activité élevée du système d'ocytocine dans le corps, et r- à une faible activité. Ces deux styles de comportement sont déterminés génétiquement mais peuvent être modifiés, au moins temporairement, en manipulant les niveaux d'ocytocine du corps.
La reproduction est la production d'une progéniture par quelque moyen que ce soit accessible au corps.
Signification biologique
La signification biologique de la reproduction et des processus associés est très diverse. Il s'agit : d'une part - de la reproduction du nombre d'espèces et de son augmentation, par opposition à mortalité naturelle, être mangé par des prédateurs et d'autres problèmes. Deuxièmement, il offre de nouvelles combinaisons génétiques et la possibilité d'apparition de nouvelles caractéristiques chez la progéniture, ce qui permet développement évolutif groupes. De plus, lors de la reproduction, le problème de l'installation dans l'espace est souvent résolu (notamment pour les espèces sédentaires), de survivre à une période de conditions défavorables (le plus souvent au stade des œufs au repos) et d'accéder à de nouvelles ressources alimentaires (disponibles uniquement pour les juvéniles ou larves, mais pas aux organismes adultes) peut être atteint).
Problèmes et adaptations.
Seuls quelques organismes, pour la plupart primitifs, sont capables de se reproduire passivement (par exemple lorsqu'ils sont déchirés). De plus, une méthode similaire (asexuée, alias multiplication végétative) ne fournit pas l'une des fonctions principales - l'apparition de nouvelles caractéristiques chez la progéniture, qui pourraient fournir du matériel pour une évolution ultérieure. Par conséquent, en règle générale, l'essentiel pour les animaux est la reproduction sexuée. Cela nécessite : le développement de systèmes organiques spéciaux pour la maturation des cellules germinales, assurant l'accouplement de ces cellules (mâles et femelles) provenant d'individus différents, fournissant à ces cellules la nutrition nécessaire au développement et à la croissance de l'embryon, et souvent aussi des soins supplémentaires. pour les jeunes jusqu'à ce qu'ils accèdent à l'indépendance.
Au moins plusieurs moments difficiles surviennent. Premièrement, les organismes sédentaires (surtout attachés) doivent résoudre d'une manière ou d'une autre le problème de la recherche d'un partenaire et de l'accouplement (à première vue, pratiquement insoluble, surtout avec une faible densité de population). Deuxièmement, les juvéniles qui apparaissent lors de la reproduction sont de toute façon très différents des adultes - ils sont plusieurs fois plus petits, ce qui nécessite le développement de nouvelles stratégies de vie (autres mécanismes d'alimentation, protection contre les prédateurs, osmorégulation, etc.). Enfin, il est nécessaire de résoudre les problèmes liés à la croissance des juvéniles, c'est-à-dire de concevoir spécialement toutes les structures, y compris les structures squelettiques, de manière à ce qu'elles puissent croître de manière plus ou moins continue, pour finalement augmenter plusieurs fois en taille. Cependant, il est clair que les animaux ont réussi à résoudre tous ces problèmes, et ce sont plutôt les mécanismes permettant de les résoudre qui diffèrent.
K- et R - stratégies de reproduction
Les stratégies de reproduction et de soin de la progéniture sont devenues le sujet de l'une des théories écologiques générales - la théorie des stratégies R et K. On pense que tous les organismes gravitent vers l’une de ces deux stratégies de reproduction. Les stratèges K (généralement de gros animaux qui dominent les habitats stables et les communautés établies, par exemple les éléphants) se reproduisent lentement et produisent une progéniture peu nombreuse mais de grande taille, entourée d'attention et de soins. Au contraire, les stratèges R (généralement de petits animaux d'habitats perturbés, par exemple les rats) se reproduisent rapidement et en grand nombre, mais prennent peu soin de leur progéniture, ce qui s'accompagne d'une forte mortalité infantile (leur mortalité adulte est également élevée) . La stratégie K est plus rentable dans des conditions où le bien-être de la population est déterminé principalement par la concurrence, et la stratégie R est plus rentable dans des conditions de forte influence de conditions difficiles. Chez l'homme, différentes stratégies se manifestent même au sein des espèces : dans les populations urbaines (surtout dans les zones économiquement pays développés) les gens se reproduisent lentement (assurant à peine la reproduction), mais investissent beaucoup d'argent dans l'entretien, l'éducation et l'éducation des enfants. Au contraire, dans les pays pauvres agricoles des tropiques, les gens se reproduisent rapidement et activement, sans avoir les moyens de s'habiller, de chausser, d'éduquer et parfois même de nourrir décemment leurs enfants, ce qui entraîne souvent une mortalité infantile élevée, mais peut aussi s'accompagner de de fortes épidémies en nombre (qui, soit dit en passant, maintiennent en partie le niveau de vie bas dans ces pays).
Cependant, toute cette théorie a été développée principalement pour les vertébrés terrestres (et en partie pour les vertébrés terrestres). plantes supérieures). Dans l’environnement des invertébrés aquatiques, des schémas légèrement différents s’opèrent. Le plus souvent (surtout dans la mer), c'est le contraire qui se produit : des organismes grands et massifs libèrent des millions d'œufs ou de larves microscopiques dispersés ; les petits hydrobiontes se dispersent d'eux-mêmes et produisent beaucoup moins de progéniture. Expliquons cela avec des exemples.
Aperçu comparatif de la reproduction de différents taxons
Algues unicellulaires. Dans chaque groupe d'algues unicellulaires, il existe deux types de reproduction : végétative et sexuée. Végétatif - division cellulaire résultant de la mitose. Lorsqu’elles disposent de ressources, les cellules d’algues unicellulaires se reproduisent principalement de manière végétative et la taille de la population augmente de façon exponentielle. Dans des conditions défavorables à la division végétative ou pour d'autres raisons, les algues subissent une reproduction sexuée (méiose), au cours de laquelle se forment des gamètes mâles et femelles, après quoi une cellule avec un « nouveau » génotype se forme. Les cycles de vie des algues unicellulaires appartenant à différents groupes phylogénétiques diffèrent. Les cycles de nombreuses algues comprennent des étapes de repos (cellules au repos, spores, kystes, etc.) pour survivre à des conditions défavorables.
Invertébrés. Le type de reproduction initial (pour les invertébrés aquatiques, principalement marins) serait le suivant. À peu près au même moment, tous les mâles et femelles adultes grandes quantités Ils balaient leurs produits reproducteurs (œufs et spermatozoïdes) directement dans l'eau, qui eux-mêmes (s'ils ont de la chance) se retrouvent dans la colonne d'eau et s'accouplent. C’est ce qu’on appelle la fécondation externe. Le corps lui-même peut être inactif ou sédentaire. Une larve planctonique microscopique se développe à partir d'un zygote fécondé, qui flotte assez longtemps dans la colonne d'eau, s'installant avec les courants, subissant diverses transformations et passant finalement à une nutrition externe (le plus souvent du phytoplancton - la larve dite planctotrophe). En grandissant et en se préparant à passer à un mode de vie adulte, la larve s'installe sur un substrat de fond approprié et acquiert les caractéristiques d'un adulte, atteint une taille macroscopique et continue de croître pendant longtemps. Ce type de reproduction et de développement permet de résoudre tous les problèmes d'installation et de compétition intraspécifique précisément grâce aux larves (et les individus adultes peuvent même être sessiles - ils n'ont pas besoin de se rencontrer directement). D'autre part, cette approche s'accompagne d'une mortalité énorme tant parmi les gamètes que parmi les larves, ce qui nécessite leur accumulation et leur libération massives, et la synchronisation de la maturation et de la libération des cellules germinales chez différents individus de la population est extrêmement importante. Ceci est réalisé par la libération de substances signal dans l'eau, qui stimulent les individus à libérer dans l'eau tous les gamètes accumulés à l'avance. En règle générale, le frai massif a lieu une fois par an et dans de nombreux organismes, une fois dans la vie. Comme il est facile de le comprendre, cette stratégie convient aux organismes relativement grands, massifs, massifs et sédentaires : polypes, éponges, mollusques, grands polychètes, échinodermes et crustacés. En général, en mer, cette option est considérée comme la plus typique.
Et aussi les petits invertébrés mobiles (cladocères et copépodes, certains petits polychètes, oligochètes, escargots) ne peuvent pas se permettre une libération massive de gamètes dans l'eau (simplement n'ayant pas assez de masse), et ont recours à la fécondation interne : ils se retrouvent et s'accouplent, après quoi la femelle porte généralement pendant un certain temps développer des œufs en eux-mêmes (réduire leur mortalité). Soit des œufs passifs protégés par une coquille spéciale, soit des larves déjà actives naissent. Les larves mènent le plus souvent un mode de vie similaire à celui des adultes ; mais sont souvent plus mobiles, ce qui permet à la population d'être mieux répartie dans l'espace. Parfois, dans ce cas, les larves d'organismes benthiques deviennent planctoniques pendant un certain temps. Souvent (par exemple, chez les oligochètes), il n'y a pas de larves du tout et les juvéniles ont une structure et un mode de vie similaires à ceux des adultes ( développement direct). Tout cela permet de générer plusieurs ordres de grandeur en moins de produits reproducteurs, réduisant ainsi les coûts de reproduction, et en même temps de se reproduire toute l'année, sans se soucier de la synchronisation du frai. Souvent, à la naissance, la larve reçoit un apport de nutriments suffisant pour mener à bien toute sa vie de migration larvaire et ne se nourrit pas du tout (larve lécithotrophe).
Dans les eaux douces, la reproduction selon le premier type (avec fécondation externe et stade larvaire planctonique long) est compliquée par des problèmes osmotiques : l'osmorégulation des gamètes flottants et des larves planctoniques s'est avérée extrêmement gênante, et la plupart des invertébrés même inférieurs utilisent la fécondation interne - et aucune larve planctonique supplémentaire. En règle générale, des œufs assez gros sont pondus - en petites quantités, mais avec un apport décent en nutriments, ce qui permet à l'organisme d'être largement lécithotrophe et d'éclore, étant déjà assez macroscopique et doté d'un système d'osmorégulation développé. C'est le chemin des vers d'eau douce, des escargots et de la plupart des crustacés. Les copépodes (comme les Cyclopes) possèdent encore une larve planctonique (nauplius), mais de durée de vie relativement courte, atteignant rapidement une apparence définitive (adulte) au cours d'une série de mues successives.
Les insectes, dans leur ensemble, sont terrestres et très mobiles précisément au stade adulte (imago), lors de leur développement. Environnement aquatique ont développé leur propre stratégie de reproduction et de cycle de vie. Ils ont laissé la fonction de dispersion (ainsi que l'accouplement et la ponte) aux individus adultes, et les larves qui vivent dans l'eau (et généralement beaucoup plus longtemps que les adultes) sont responsables de la nutrition, de la croissance (et de l'accumulation de nutriments dans le corps), ainsi que la survie dans l'eau lors des saisons défavorables à la vie terrestre (principalement l'hiver). Les larves d'insectes, même lorsqu'elles éclosent d'œufs, sont macroscopiques, capables de se nourrir de manière indépendante et disposent d'un système d'osmorégulation en eau douce (et parfois saumâtre) tout à fait parfait. Il est intéressant de noter que les adultes de certains groupes (éphémères, caddisflies, chironomides, certaines mouches de pierre) ne se nourrissent pas du tout et vivent très peu de temps, et leur vol synchronisé depuis les plans d'eau est utilisé pour une reproduction réussie. Ainsi, les insectes adultes chez les non-insectes sont fonctionnellement équivalents aux produits reproducteurs (gamètes) de nombreux invertébrés marins.
Chez certains groupes d'invertébrés (plus souvent dans les eaux douces que dans les eaux marines), il est répandu hermaphrodisme - lorsque les organes reproducteurs et les gamètes mâles et femelles se forment chez chaque individu. Par exemple, tous les escargots pulmonaires (Pulmonata), les oligochètes et les balanes sont hermaphrodites. Lors de l'accouplement, un organisme peut agir à la fois comme un mâle et une femelle, et souvent comme les deux à la fois (on observe alors une fécondation mutuelle). La signification biologique de l’hermaphrodisme (c’est-à-dire la culture d’un double ensemble d’organes dans chaque corps) n’est pas tout à fait claire. Parfois (mais apparemment rarement) l'autofécondation se produit - cela viole en partie l'idée même de reproduction sexuée (puisque l'organisme se croise avec lui-même), mais elle permet à un seul individu de donner naissance à une nouvelle population dans un nouvel endroit. .
L'hermaphrodisme chez les animaux est encore moins fréquent reproduction asexuée, dans lequel les mères se clonent elles-mêmes, produisant des femelles génétiquement identiques. Cette situation est particulièrement typique lors des périodes d'épidémies de petits invertébrés d'eau douce - en particulier les daphnies et les rotifères dans période estivale. Dans tous les cas, il s’agit d’une mesure temporaire, qui tôt ou tard (généralement à l’automne) cède la place à une reproduction sexuée normale. Cependant, chez les protozoaires unicellulaires (comme chez les plantes), la reproduction asexuée est la chose la plus courante ; c'est grâce à elle que se produit la principale reproduction des espèces.
Poisson. En règle générale, les poissons ont une fécondation externe, mais celle-ci est réalisée avec rencontre personnelle individus parentaux (la femelle pond ses œufs et le mâle lui donne immédiatement du lait). En conséquence, les poissons pondent des œufs, généralement assez petits et en grande quantité. Le nombre d'œufs est en moyenne de plusieurs milliers, mais varie considérablement selon les différents types: de 10-30 pièces (pour les épinoches) à 10-100 millions (pour le thon, le cabillaud et bien d'autres gros poisson de mer). Dans le même temps, les œufs contiennent un certain apport de nutriments, ce qui leur permet d'éclore pour donner des alevins entièrement formés, capables de nager et de se nourrir. Les alevins ne maîtrisent pas de nouveaux environnements, mais ils captent des spectres alimentaires habituellement inaccessibles aux poissons adultes : ils peuvent se nourrir de zooplancton et de méiobenthos. Certes, il n'est pas clair si les poissons dans leur ensemble bénéficient grandement de cette circonstance ou s'il s'agit d'une mesure nécessaire (puisque les alevins ne peuvent pas manger autre chose en raison de leur petite taille).
Certaines espèces de poissons ont cependant des formes de reproduction et de protection de la progéniture assez bizarres. Les plus connus sont les poissons migrateurs qui changent d'habitat pour se reproduire. Le saumon et l'esturgeon vivent dans les mers à l'âge adulte et frayent dans les rivières (d'où ils sont originaires), et leurs petits, adaptés à l'osmorégulation de l'eau douce, restent dans les rivières pendant un certain temps et descendent ensuite seulement dans la mer. Dans le même temps, les saumons font des miracles d'héroïsme en surmontant les rapides des rivières de la taïga de montagne ; et peu de temps après le frai, ils meurent - directement dans les rivières, augmentant considérablement leur saprobité. Il s'avère qu'il s'agit d'un moyen unique de saturer l'habitat des juvéniles en matière organique. Une autre question est de savoir quelle est son efficacité.
L'anguille, au contraire, nage des rivières jusqu'à la mer des Sargasses pour se reproduire, et sa population européenne surmonte à cet effet (en aval) la quasi-totalité de la population. océan Atlantique. Les jeunes (toujours en aval, mais de manière différente) retournent dans les rivières européennes. Cela ne semble pas avoir beaucoup de sens. On pense que les migrations ultra-longues des anguilles reflètent la dérive des continents, au cours de laquelle l'Atlantique s'étend progressivement, et les anguilles doivent nager de plus en plus loin dans leur mer d'origine tous les millions d'années.
Certains groupes de poissons ont adopté une stratégie K explicite, principalement par viviparité. Dans ce cas, la fécondation est interne, le nombre de descendants est beaucoup plus petit, mais au moment où ils entrent dans l'eau, ils sont plus grands et plus viables. L’exemple le plus célèbre est celui des poissons d’aquarium vivipares Peciliidae (guppys et autres platies). Tous les aquariophiles savent qu’il est beaucoup plus facile de les élever que n’importe quel autre poisson. Les requins, par exemple, font la même chose : ils pondent très peu d'œufs (généralement 5 à 30), mais de très gros œufs - le requin baleine mesure jusqu'à 60 cm (!) de diamètre, ce qui leur permet d'éclore pour donner de très gros poissons. .
Amphibiens. Les amphibiens ont une fécondation interne et pondent des œufs assez gros – et toujours dans l’eau. Comme les insectes, la plupart des amphibiens sont amphibiotiques, c'est-à-dire qu'ils se reproduisent dans l'eau et possèdent des larves ressemblant à des poissons aquatiques (têtards), bien que les animaux adultes vivent sur terre la majeure partie de leur vie. En général, on peut aussi parler ici de l'interception de nouveaux habitats et ressources alimentaires par les têtards - cela est généralement vrai, mais reflète en fait l'infériorité globale de l'ensemble de la classe - les amphibiens ne savent tout simplement pas comment faire autrement.
Crabes et prendre soin d'une femme. Chez de nombreux crustacés, notamment supérieurs, les adultes sont si bien protégés par une coquille chitineuse qu'ils ne peuvent s'accoupler qu'immédiatement après la mue de la femelle. Ainsi, un mâle prêt à s'accoupler doit non seulement trouver une femelle de sa propre espèce, mais aussi attendre qu'elle mue, ce qui peut arriver, par exemple, en quelques semaines. De plus, vous devez attendre à proximité et ne pas chercher une femelle en mue - car pendant et après la mue, les animaux deviennent extrêmement vulnérables et tentent de muer dans des abris fiables (où ils sont difficiles à trouver). Ainsi, par exemple, parmi les crabes du Kamtchatka, les mâles adultes rassemblent plusieurs femelles autour d'eux (un harem) et les « broutent », s'accouplent avec celles qui ont mué et les protègent d'être mangées (principalement par d'autres femelles de leur propre harem). De tels soins vitaux à l’égard de la femelle n’ont pas grand-chose à voir avec la « parade nuptiale » frivole des vertébrés. La situation est encore compliquée par le fait que si le mâle lui-même mue, il peut aussi être immédiatement mangé par ses femelles, donc pour muer, il est obligé de quitter son harem et de se cacher soigneusement.
Harpacticides et pédophilie. Ces petits copépodes ont un faible dimorphisme sexuel et leurs différences interspécifiques sont faibles ; et les changements liés à l'âge (des stades copépodites des juvéniles aux stades sexuellement matures) sont peu perceptibles. Mais l’instinct sexuel des mâles est très fort. Par conséquent, un mâle prêt à s'accoupler, fouillant dans le limon du fond à la recherche d'un partenaire sexuel, ne fait preuve d'aucune sélectivité et s'accouple avec presque n'importe qui - avec une femelle de sa propre espèce (s'il a de la chance !), ou avec un mâle, soit avec un copépodite (c'est-à-dire un jeune individu), soit avec un crustacé d'une espèce complètement différente. Parfois, leur activité similaire est confondue avec une tentative de manger leur partenaire, mais il s'agit ici d'une tentative de copulation. En état d'accouplement, les crustacés nagent assez longtemps, et si un mâle est également dans la position de la femelle, il peut, entre-temps, également attraper un partenaire pour l'accouplement ; Parfois, cela produit des chaînes d’individus assez longues, dont seulement quelques-uns s’accouplent.
Escargots et accouplement en groupe. Les escargots d'eau douce pulmonaires sont hermaphrodites et, chez certains d'entre eux, la détermination du sexe lors de l'accouplement est directement déterminée par la position de l'animal lui-même - approximativement selon le principe « celui qui est au sommet est le mâle ». Par exemple, les calices des rivières ( Ancylus fluviatilis) pour s'accoupler, ils rampent simplement les uns sur les autres, puis pendent leurs organes copulatoires. Cette situation n’empêche pas une autre tasse de ramper sur le dessus et de copuler avec celle du dessous, et ainsi de suite. En conséquence, une pile d'individus copulants peut se former, dont le plus bas agit uniquement comme une femelle, et le plus élevé comme un mâle, et tous les autres travaillent avec les deux systèmes organiques (contrairement aux harpacticides stupides, qui ne peuvent qu'imiter une situation similaire). Ensuite, tout le monde rampe et pond des œufs ensemble.
Bonellia et la détermination du sexe par le destin. Chez l'échiuridé marin sessile Bonellia, la larve planctonique, entrant en nage libre, n'a pas encore de sexe spécifique, mais a déjà non seulement une dispersion, mais aussi une tâche sexuelle - la recherche d'une femelle. Si la larve parvient à trouver une bonellia femelle adulte, elle la pénètre et se développe en un mâle (qui vit ensuite à l'intérieur de la femelle pour le reste de sa vie, la fécondant). Si aucune femelle n’est trouvée, la larve finit par s’installer au fond et devient elle-même une femelle.
"... deux scientifiques américains, Robert MacArthur et Édouard Wilson, créé Théorie RK sélection. La théorie de deux différentes stratégies reproduction des êtres vivants.
La théorie des deux stratégies s'est avérée si réussie qu'elle est utilisée dans un certain nombre de sciences, reconnue par presque tout le monde et incluse dans les manuels et les supports pédagogiques.
La stratégie R est la naissance par unité de temps autant que possible plus petits.
Chacun d'eux ne peut pratiquement pas être soigné et chaque ourson n'a pas beaucoup de chances de survivre. Une mouche pond 5 millions d'œufs - et s'inquiète-t-elle vraiment du sort de ces 5 millions de futures petites mouches ? Des centaines de milliers et des millions d’insectes, de crustacés et de mollusques pondent des œufs. Les poissons qui pondent « seulement » des dizaines de milliers d’œufs, en particulier les grenouilles qui en pondent des milliers, sont tout simplement des parents idéaux par rapport aux créatures plus simples. Bien sûr, ils ne se soucient pas non plus de leur progéniture, mais ces animaux plus complexes sont obligés de pondre des œufs plus complexes et plus gros - et ainsi de pondre moins de ces œufs. Certaines espèces de poissons tentent déjà de protéger leurs poissons éclos : ils leur construisent des nids et attaquent les prédateurs émergents. Certaines espèces gardent même les alevins dans leur propre bouche, où ils s'échappent en cas de danger.
Ce sont déjà des éléments de la stratégie K : la naissance d'un petit nombre de petits, dont chacun est important et précieux. Plus l'espèce est complexe, plus chaque vie individuelle a de la valeur pour elle, plus moins de petits meurt entre la naissance et la mort. Plus c'est simple Être vivant, moins il a besoin d’être instruit et préparé à la vie, plus vite il devient adulte.
Une souris peut donner naissance à dix petites souris trois fois par an. Donner naissance à une souris est très simple et les bébés deviennent adultes en trois semaines. Ils peuvent déjà prendre soin d'eux-mêmes, la mère les met à la porte et est prête à en donner naissance à de nouveaux. Si les petites souris ne meurent pas, le monde sera bientôt rempli de hordes de souris adultes. Les animaux plus complexes - éléphants, chimpanzés, élans, bisons - ont moins de petits et meurent moins souvent.
Mais même chez les grands animaux complexes, la norme physiologique est la mortalité. 60-70% les nouveau-nés. Une femelle chimpanzé et un éléphant donnent naissance à 10 à 15 fois au cours de leur vie. 7, 10 voire 12 de ces bébés mourront avant de devenir adultes. Les mêmes 2 ou 3 petits nécessaires à la reproduction de l'espèce grandiront et donneront naissance à une tribu.
Après des catastrophes dues à des explosions volcaniques, après un tsunami, de nouvelles îles et côtes sont « capturées » par des êtres vivants avec une stratégie R. Mais bientôt, des animaux plus grands et plus complexes dotés d’une stratégie K commencent à dominer. À bien des égards, l’évolution est une lutte non pas pour la survie, mais pour la domination. »
Burovsky A.M., Phénomène cérébral. Les secrets de 100 milliards de neurones, M., « Yauza » ; "Eksmo", 2010, p. 77-79.