លំយោលនៃប្រព័ន្ធសត្វមំសាសី (គំរូ Lotka-Voltaire) ។ វគ្គសិក្សា៖ ការស្រាវជ្រាវគុណភាពនៃគំរូ predator-prey គំរូគណិតវិទ្យានៃប្រព័ន្ធ prey-predator
គំរូនៃអន្តរកម្មនៃប្រភេទពីរ
សម្មតិកម្មរបស់ Volterra ។ អាណាឡូកជាមួយគីមីវិទ្យា។ គំរូ Volterra នៃអន្តរកម្ម។ ចំណាត់ថ្នាក់នៃប្រភេទអន្តរកម្ម ការប្រកួតប្រជែង។ Predator - សត្វព្រៃ។ គំរូទូទៅនៃអន្តរកម្មប្រភេទ . ម៉ូដែល Kolmogorov ។ គំរូរបស់ MacArthur នៃអន្តរកម្មរវាងសត្វល្អិតពីរប្រភេទ។ ប៉ារ៉ាម៉ែត្រ និងរូបភាពដំណាក់កាលនៃប្រព័ន្ធ Bazykin ។
ស្ថាបនិកនៃទ្រឹស្តីគណិតវិទ្យាសម័យទំនើបនៃចំនួនប្រជាជនត្រូវបានគេចាត់ទុកថាជាគណិតវិទូជនជាតិអ៊ីតាលី Vito Volterra ដែលបានបង្កើតទ្រឹស្តីគណិតវិទ្យានៃសហគមន៍ជីវសាស្រ្ត ដែលជាឧបករណ៍នៃសមីការឌីផេរ៉ង់ស្យែល និងអាំងតេក្រាលឌីផេរ៉ង់ស្យែល។(Vito Volterra. Lecons sur la Theorie Mathematique de la Lutte pour la Vie. Paris, ១៩៣១)។ ក្នុងទសវត្សរ៍ជាបន្តបន្ទាប់ សក្ដានុពលចំនួនប្រជាជនបានអភិវឌ្ឍយ៉ាងសំខាន់ស្របតាមគំនិតដែលបានបង្ហាញនៅក្នុងសៀវភៅនេះ។ ការបកប្រែជាភាសារុស្សីនៃសៀវភៅរបស់ Volterra ត្រូវបានបោះពុម្ពនៅឆ្នាំ 1976 ក្រោមចំណងជើងថា "ទ្រឹស្តីគណិតវិទ្យានៃការតស៊ូដើម្បីអត្ថិភាព" ជាមួយនឹងពាក្យក្រោយដោយ Yu.M. Svirezhev ដែលពិនិត្យមើលប្រវត្តិសាស្រ្តនៃការអភិវឌ្ឍន៍នៃបរិស្ថានវិទ្យាគណិតវិទ្យាក្នុងកំឡុងឆ្នាំ 1931-1976 ។
សៀវភៅរបស់ Volterra ត្រូវបានសរសេរតាមរបៀបដែលសៀវភៅគណិតវិទ្យាត្រូវបានសរសេរ។ ដំបូងវាបង្កើតការសន្មត់មួយចំនួនអំពីវត្ថុគណិតវិទ្យាដែលគេសន្មត់ថាត្រូវសិក្សា ហើយបន្ទាប់មកធ្វើការសិក្សាគណិតវិទ្យាអំពីលក្ខណៈសម្បត្តិរបស់វត្ថុទាំងនេះ។
ប្រព័ន្ធដែលបានសិក្សាដោយ Volterra មានពីរប្រភេទ ឬច្រើន។ ក្នុងករណីខ្លះការផ្គត់ផ្គង់អាហារដែលបានប្រើត្រូវបានពិចារណា។ សមីការដែលពិពណ៌នាអំពីអន្តរកម្មនៃប្រភេទទាំងនេះគឺផ្អែកលើគោលគំនិតខាងក្រោម។
សម្មតិកម្មរបស់ Volterra
1. អាហារអាចរកបានក្នុងបរិមាណមិនកំណត់ ឬការផ្គត់ផ្គង់របស់វាត្រូវបានគ្រប់គ្រងយ៉ាងតឹងរ៉ឹងតាមពេលវេលា។
2. បុគ្គលនៃប្រភេទនីមួយៗស្លាប់តាមរបៀបដែលសមាមាត្រថេរនៃបុគ្គលដែលមានស្រាប់ស្លាប់ក្នុងមួយឯកតាពេលវេលា។
3. ប្រភេទសត្វព្រៃស៊ីសាច់ជនរងគ្រោះ ហើយក្នុងមួយឯកតាចំនួននៃជនរងគ្រោះដែលបរិភោគគឺតែងតែសមាមាត្រទៅនឹងប្រូបាប៊ីលីតេនៃការជួបបុគ្គលនៃប្រភេទទាំងពីរនេះ ពោលគឺឧ។ ផលិតផលនៃចំនួនសត្វមំសាសី និងចំនួនសត្វព្រៃ។
4. ប្រសិនបើមានអាហារក្នុងបរិមាណកំណត់ និងប្រភេទជាច្រើនដែលអាចទទួលទានបាន នោះចំណែកនៃអាហារដែលប្រើប្រាស់ដោយប្រភេទសត្វក្នុងមួយឯកតាពេលវេលាគឺសមាមាត្រទៅនឹងចំនួនបុគ្គលនៃប្រភេទសត្វនេះ ដោយយកដោយមេគុណជាក់លាក់មួយអាស្រ័យលើ ប្រភេទ (គំរូនៃការប្រកួតប្រជែងគ្នា) ។
5. ប្រសិនបើប្រភេទសត្វចិញ្ចឹមលើអាហារដែលមានក្នុងបរិមាណមិនកំណត់ នោះការកើនឡើងនៃចំនួនប្រភេទក្នុងមួយឯកតាពេលវេលាគឺសមាមាត្រទៅនឹងចំនួនប្រភេទសត្វ។
6. ប្រសិនបើប្រភេទសត្វចិញ្ចឹមលើអាហារដែលមានក្នុងបរិមាណកំណត់ នោះការបន្តពូជរបស់វាត្រូវបានគ្រប់គ្រងដោយអត្រានៃការប្រើប្រាស់អាហារ ពោលគឺឧ។ ក្នុងមួយឯកតាពេល ការកើនឡើងគឺសមាមាត្រទៅនឹងបរិមាណអាហារដែលបរិភោគ។
អាណាឡូកជាមួយគីមីវិទ្យា
សម្មតិកម្មទាំងនេះមានភាពស្រដៀងគ្នាយ៉ាងជិតស្និទ្ធជាមួយ kinetics គីមី។ នៅក្នុងសមីការនៃសមីការនៃចំនួនប្រជាជន ដូចជានៅក្នុងសមីការនៃ kinetics គីមី "គោលការណ៍ប៉ះទង្គិច" ត្រូវបានប្រើនៅពេលដែលអត្រាប្រតិកម្មគឺសមាមាត្រទៅនឹងផលិតផលនៃការប្រមូលផ្តុំនៃសមាសធាតុប្រតិកម្ម។
ជាការពិតណាស់យោងទៅតាមសម្មតិកម្មរបស់ Volterra ល្បឿនដំណើរការ ការផុតពូជនៃប្រភេទសត្វនីមួយៗគឺសមាមាត្រទៅនឹងចំនួននៃប្រភេទសត្វ។ នៅក្នុង kinetics គីមី នេះត្រូវគ្នាទៅនឹងប្រតិកម្ម monomolecular នៃការ decomposition នៃសារធាតុជាក់លាក់មួយ ហើយនៅក្នុងគំរូគណិតវិទ្យា វាត្រូវគ្នាទៅនឹងពាក្យលីនេអ៊ែរអវិជ្ជមាននៅជ្រុងខាងស្តាំនៃសមីការ។
យោងតាមគោលគំនិតនៃ kinetics គីមី អត្រានៃប្រតិកម្ម bimolecular នៃអន្តរកម្មរវាងសារធាតុទាំងពីរគឺសមាមាត្រទៅនឹងប្រូបាប៊ីលីតេនៃការប៉ះទង្គិចនៃសារធាតុទាំងនេះ i.e. ផលិតផលនៃការផ្តោតអារម្មណ៍របស់ពួកគេ។ នៅក្នុងវិធីដូចគ្នានេះ អនុលោមតាមសម្មតិកម្មរបស់ Volterra អត្រានៃការបន្តពូជរបស់សត្វមំសាសី (ការស្លាប់របស់សត្វព្រៃ) គឺសមាមាត្រទៅនឹងប្រូបាប៊ីលីតេនៃការជួបគ្នារវាងសត្វមំសាសី និងបុគ្គលសត្វ ពោលគឺឧ។ ផលិតផលនៃលេខរបស់ពួកគេ។ ក្នុងករណីទាំងពីរនេះ ពាក្យ bilinear លេចឡើងនៅក្នុងប្រព័ន្ធគំរូនៅខាងស្តាំដៃនៃសមីការដែលត្រូវគ្នា។
ជាចុងក្រោយ លក្ខខណ្ឌវិជ្ជមានលីនេអ៊ែរនៅខាងស្តាំដៃនៃសមីការ Volterra ដែលត្រូវគ្នាទៅនឹងកំណើនប្រជាជនក្រោមលក្ខខណ្ឌគ្មានដែនកំណត់ ត្រូវគ្នាទៅនឹងលក្ខខណ្ឌ autocatalytic ប្រតិកម្មគីមី. ភាពស្រដៀងគ្នានៃសមីការនេះនៅក្នុងគំរូគីមី និងបរិស្ថានអនុញ្ញាតឱ្យយើងអនុវត្តវិធីសាស្រ្តស្រាវជ្រាវដូចគ្នាសម្រាប់គំរូគណិតវិទ្យានៃ kinetics ចំនួនប្រជាជន ដូចជាសម្រាប់ប្រព័ន្ធនៃប្រតិកម្មគីមី។
ការបែងចែកប្រភេទនៃអន្តរកម្ម
អនុលោមតាមសម្មតិកម្មរបស់ Volterra អន្តរកម្មនៃប្រភេទសត្វពីរដែលជាលេខ x 1 និង x 2 អាចត្រូវបានពិពណ៌នាដោយសមីការ:
(9.1)
នេះគឺជាប៉ារ៉ាម៉ែត្រ ក ខ្ញុំ - អត្រាកំណើនផ្ទាល់ខ្លួនរបស់ប្រភេទសត្វ គ ខ្ញុំ- អថេរនៃការកំណត់ដោយខ្លួនឯងនៃលេខ (ការប្រកួតប្រជែងមិនជាក់លាក់), b ij- អថេរអន្តរកម្មប្រភេទ, (ខ្ញុំ, j=១,២)។ សញ្ញានៃមេគុណទាំងនេះកំណត់ប្រភេទនៃអន្តរកម្ម។
នៅក្នុងអក្សរសិល្ប៍ជីវសាស្រ្ត អន្តរកម្មជាធម្មតាត្រូវបានចាត់ថ្នាក់តាមយន្តការដែលពាក់ព័ន្ធ។ ភាពចម្រុះនៅទីនេះគឺធំសម្បើម៖ អន្តរកម្ម trophic ផ្សេងៗ អន្តរកម្មគីមីដែលមានស្រាប់រវាងបាក់តេរី និងសារាយ planktonic អន្តរកម្មនៃផ្សិតជាមួយសារពាង្គកាយផ្សេងទៀត ការបន្តពូជនៃសារពាង្គកាយរុក្ខជាតិ ដែលត្រូវបានផ្សារភ្ជាប់ជាពិសេសជាមួយនឹងការប្រកួតប្រជែងសម្រាប់ ពន្លឺព្រះអាទិត្យនិងជាមួយនឹងការវិវត្តនៃដី។ល។ ការចាត់ថ្នាក់នេះហាក់ដូចជាធំ។
អ៊ី . Odum ដោយគិតគូរពីគំរូដែលស្នើឡើងដោយ V. Volterra បានស្នើការចាត់ថ្នាក់មិនមែនដោយយន្តការទេ ប៉ុន្តែដោយលទ្ធផល។ យោងតាមការចាត់ថ្នាក់នេះ ទំនាក់ទំនងគួរតែត្រូវបានវាយតម្លៃថាជាវិជ្ជមាន អវិជ្ជមាន ឬអព្យាក្រឹត អាស្រ័យលើថាតើភាពសម្បូរបែបនៃប្រភេទសត្វមួយកើនឡើង ថយចុះ ឬនៅតែមិនផ្លាស់ប្តូរនៅក្នុងវត្តមាននៃប្រភេទសត្វមួយផ្សេងទៀត។ បន្ទាប់មក ប្រភេទសំខាន់ៗនៃអន្តរកម្មអាចត្រូវបានបង្ហាញជាទម្រង់តារាង។
ប្រភេទនៃអន្តរកម្មនៃប្រភេទសត្វ
|
ស៊ីមប៊ីស៊ីស |
ខ 12 ,ខ 21 >0 |
||
|
CommENSALISM |
ខ 12 ,>0, ខ 21 =0 |
||
|
PREDATOR-ជនរងគ្រោះ |
ខ 12 ,>0, ខ 21 <0 |
||
|
អាម៉ែនសាលីស |
ខ 12 ,=0, ខ 21 <0 |
||
|
ការប្រកួតប្រជែង |
ខ 12 , ខ 21 <0 |
||
|
អព្យាក្រឹតភាព |
ខ 12 , ខ 21 =0 |
ជួរចុងក្រោយបង្ហាញពីសញ្ញានៃមេគុណអន្តរកម្មពីប្រព័ន្ធ (9.1)
សូមក្រឡេកមើលប្រភេទសំខាន់ៗនៃអន្តរកម្ម
សមភាពប្រកួតប្រជែង៖
ដូចដែលយើងបានឃើញនៅក្នុងមេរៀនទី 6 សមីការនៃការប្រកួតប្រជែងគឺ:
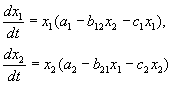 (9.2)
(9.2)
ដំណោះស្រាយប្រព័ន្ធស្ថានី៖
(1).
![]()
ប្រភពដើមនៃកូអរដោណេ សម្រាប់ប៉ារ៉ាម៉ែត្រប្រព័ន្ធណាមួយ គឺជាថ្នាំងដែលមិនស្ថិតស្ថេរ។
(2).
![]() (9.3)
(9.3)
គ ស្ថានភាពស្ថានី (9.3) គឺជាកន្លែងមួយនៅ ក 1 > ខ 12 / ជាមួយ 2 និង
ថ្នាំងមានស្ថេរភាពនៅ ក 1 12 /s ២ . លក្ខខណ្ឌនេះមានន័យថាប្រភេទសត្វមួយប្រភេទបានផុតពូជ ប្រសិនបើអត្រាកំណើនរបស់វាមានចំនួនតិចជាងតម្លៃសំខាន់ជាក់លាក់។
(3).
![]() (9.4)
(9.4)
គ ដំណោះស្រាយស្ថានី (9.4)¾ saddle នៅ ក 2 > ខ 21 / គ 1 និងថ្នាំងដែលមានស្ថេរភាពនៅ ក 2< ខ 21 / គ 1
(4).
![]() (9.5)
(9.5)
ស្ថានភាពស្ថានី (9.5) កំណត់លក្ខណៈនៃការរួមរស់នៃប្រភេទប្រកួតប្រជែងពីរ ហើយតំណាងឱ្យថ្នាំងដែលមានស្ថេរភាព ប្រសិនបើទំនាក់ទំនងត្រូវបានពេញចិត្ត៖
![]()
នេះបង្ហាញពីវិសមភាព៖
ខ 12
ខ 21
អនុញ្ញាតឱ្យយើងបង្កើតលក្ខខណ្ឌសម្រាប់ការរួមរស់នៃប្រភេទសត្វ:
ផលិតផលនៃមេគុណនៃអន្តរកម្មប្រជាជនគឺតិចជាងផលិតផលនៃមេគុណនៅក្នុងអន្តរកម្មប្រជាជន។
ពិតហើយ សូមឲ្យអត្រាកំណើនធម្មជាតិនៃប្រភេទទាំងពីរនេះស្ថិតក្រោមការពិចារណាក 1 , ក 2 គឺដូចគ្នា។ បន្ទាប់មកលក្ខខណ្ឌចាំបាច់សម្រាប់ស្ថេរភាពនឹងមាន
គ 2 > ខ 12 , គ 1 > ខ 21 .
វិសមភាពទាំងនេះបង្ហាញថា ការកើនឡើងនៃទំហំនៃដៃគូប្រកួតប្រជែងមួយរារាំងកំណើនរបស់ខ្លួនច្រើនជាងកំណើននៃដៃគូប្រកួតប្រជែងផ្សេងទៀត។ ប្រសិនបើចំនួននៃប្រភេទទាំងពីរមានកម្រិត មួយផ្នែក ឬទាំងស្រុងដោយធនធានផ្សេងគ្នា វិសមភាពខាងលើមានសុពលភាព។ ប្រសិនបើប្រភេទសត្វទាំងពីរមានតម្រូវការដូចគ្នា នោះមួយក្នុងចំណោមពួកវានឹងអាចសម្រេចបានជាង ហើយនឹងផ្លាស់ប្តូរដៃគូប្រកួតប្រជែងរបស់វា។
អាកប្បកិរិយានៃគន្លងដំណាក់កាលនៃប្រព័ន្ធផ្តល់នូវគំនិតច្បាស់លាស់អំពីលទ្ធផលដែលអាចកើតមាននៃការប្រកួតប្រជែង។ ចូរយើងយកផ្នែកខាងស្តាំនៃសមីការនៃប្រព័ន្ធ (9.2) ទៅសូន្យ៖
x 1 (ក១–គ ១ x 1 – ខ 12 x 2) = 0 (dx 1 /dt = 0),
x 2 (ក 2 –ខ 21 x 1 – គ 2 x 2) = 0 (dx 2 /dt = 0),
ក្នុងករណីនេះ យើងទទួលបានសមីការសម្រាប់ isoclines សំខាន់នៃប្រព័ន្ធ
x២ = − ខ ២១ x 1 / គ 2 +ក២/គ ២, x 2 = 0
- សមីការនៃ isoclines នៃតង់សង់បញ្ឈរ។
x២ = – គ ១ x 1 / ខ ១២ + ក 1 / ខ 12 , x 1 = 0
- សមីការនៃ isoclines នៃតង់សង់បញ្ឈរ។ ចំនុចប្រសព្វជាគូនៃ isoclines នៃប្រព័ន្ធតង់សង់បញ្ឈរ និងផ្ដេក តំណាងឱ្យដំណោះស្រាយស្ថានីនៃប្រព័ន្ធសមីការ (9.2.) និងកូអរដោនេរបស់វា ![]() គឺជាចំនួនថេរនៃប្រភេទសត្វដែលប្រកួតប្រជែង។
គឺជាចំនួនថេរនៃប្រភេទសត្វដែលប្រកួតប្រជែង។
ទីតាំងដែលអាចកើតមាននៃ isoclines សំខាន់ៗនៅក្នុងប្រព័ន្ធ (9.2) ត្រូវបានបង្ហាញក្នុងរូប 9.1។ អង្ករ។ ៩.១កទាក់ទងទៅនឹងការរស់រានមានជីវិតនៃប្រភេទសត្វx 1, រូបភព។ ៩.១ ខ- ការរស់រានមានជីវិតនៃប្រភេទសត្វx 2, រូបភព។ ៩.១ វ- ការរួមរស់នៃប្រភេទសត្វនៅពេលដែលលក្ខខណ្ឌ (9.6) ត្រូវបានពេញចិត្ត។ រូបភាព 9.1ជីបង្ហាញពីប្រព័ន្ធកេះ។ នៅទីនេះលទ្ធផលនៃការប្រកួតប្រជែងគឺអាស្រ័យលើលក្ខខណ្ឌដំបូង។ ស្ថានភាពមិនសូន្យ (9.5) សម្រាប់ប្រភេទទាំងពីរគឺមិនស្ថិតស្ថេរ។ នេះគឺជាសោរដែលឆ្លងកាត់ដោយបំបែកតំបន់នៃការរស់រានមានជីវិតនៃប្រភេទនីមួយៗ។

អង្ករ។ ៩.១.ទីតាំងនៃ isoclines សំខាន់នៅលើបញ្ឈរដំណាក់កាលនៃប្រព័ន្ធ Volterra នៃការប្រកួតប្រជែងនៃពីរប្រភេទ (9.2) ជាមួយនឹងសមាមាត្រផ្សេងគ្នានៃប៉ារ៉ាម៉ែត្រ។ ការពន្យល់នៅក្នុងអត្ថបទ។
ដើម្បីសិក្សាអំពីការប្រកួតប្រជែងប្រភេទសត្វ ការពិសោធន៍ត្រូវបានធ្វើឡើងលើពពួកពពួកពពួកសត្វ។ ជាធម្មតា ប្រភេទសត្វដែលទាក់ទងគ្នាយ៉ាងជិតស្និទ្ធចំនួនពីរត្រូវបានជ្រើសរើស និងដាំដុះរួមគ្នា និងដាច់ដោយឡែកពីគ្នាក្រោមលក្ខខណ្ឌគ្រប់គ្រងយ៉ាងតឹងរ៉ឹង។ នៅចន្លោះពេលជាក់លាក់ ជំរឿនពេញលេញ ឬជ្រើសរើសនៃចំនួនប្រជាជនត្រូវបានអនុវត្ត។ ទិន្នន័យពីការពិសោធន៍ចម្លងជាច្រើនត្រូវបានកត់ត្រា និងវិភាគ។ ការសិក្សាត្រូវបានអនុវត្តលើប្រូតូហ្សូអា (ជាពិសេស ciliates) ប្រភេទសត្វជាច្រើននៃពពួកពពួក Tribolium, drosophila និង crustaceans ទឹកសាប (daphnia) ។ ការពិសោធន៍ជាច្រើនត្រូវបានអនុវត្តលើចំនួនអតិសុខុមប្រាណ (សូមមើលមេរៀនទី 11) ។ ការពិសោធន៍ក៏ត្រូវបានអនុវត្តនៅក្នុងធម្មជាតិ រួមទាំងនៅលើ planarians (Reynolds) ស្រមោចពីរប្រភេទ (Pontin) ជាដើម។ នៅក្នុងរូបភព។ ៩.២. ពិពណ៌នាអំពីខ្សែកោងកំណើននៃ diatoms ដោយប្រើធនធានដូចគ្នា (កាន់កាប់ផ្នែកអេកូឡូស៊ីដូចគ្នា)។ នៅពេលដាំដុះក្នុងវប្បធម៍ទោល។ Asterionella Formosa ឈានដល់កម្រិតដង់ស៊ីតេថេរ និងរក្សាកំហាប់នៃធនធាន (ស៊ីលីត) នៅកម្រិតទាបឥតឈប់ឈរ។ ខ.ពេលដាំដុះក្នុងលក្ខណៈមនោគមវិជ្ជា Synedrauina មានឥរិយាបទស្រដៀងគ្នា និងរក្សាកំហាប់ស៊ីលីកេតនៅកម្រិតទាបជាង។ ខ.កំឡុងពេលដាំដុះរួមគ្នា (ស្ទួន) Synedrauina ផ្លាស់ទីលំនៅ Asterionella formosa ។ ជាក់ស្តែង Synedra

អង្ករ។ ៩.២.ការប្រកួតប្រជែងនៅក្នុង diatoms ។ ក -នៅពេលដាំដុះក្នុងពហុវប្បធម៍ Asterionella Formosa ឈានដល់កម្រិតដង់ស៊ីតេថេរ និងរក្សាកំហាប់នៃធនធាន (ស៊ីលីត) នៅកម្រិតទាបឥតឈប់ឈរ។ ខ -នៅពេលដាំដុះក្នុងពហុវប្បធម៍ Synedrauina មានឥរិយាបទស្រដៀងគ្នា និងរក្សាកំហាប់ស៊ីលីតនៅកម្រិតទាបជាង។ វី -ជាមួយនឹងការដាំដុះរួមគ្នា (ស្ទួន) Synedruina ផ្លាស់ទីលំនៅ Asterionella formosa ។ ជាក់ស្តែង Synedra ឈ្នះការប្រកួតប្រជែងដោយសារតែសមត្ថភាពរបស់វាក្នុងការប្រើប្រាស់ស្រទាប់ខាងក្រោមបានពេញលេញ (សូមមើលផងដែរនូវការបង្រៀន 11) ។
ការពិសោធន៍លើការសិក្សាការប្រកួតប្រជែងដោយ G. Gause ត្រូវបានគេស្គាល់យ៉ាងទូលំទូលាយ ដោយបង្ហាញពីការរស់រានមានជីវិតនៃប្រភេទសត្វដែលប្រកួតប្រជែងមួយ ហើយអនុញ្ញាតឱ្យគាត់បង្កើត "ច្បាប់នៃការមិនរាប់បញ្ចូលការប្រកួតប្រជែង" ។ ច្បាប់ចែងថាមានតែប្រភេទសត្វមួយប៉ុណ្ណោះដែលអាចមាននៅក្នុងផ្នែកអេកូឡូស៊ីមួយ។ នៅក្នុងរូបភព។ ៩.៣. លទ្ធផលនៃការពិសោធន៍របស់ Gause ត្រូវបានបង្ហាញសម្រាប់ប្រភេទ Parametium ពីរប្រភេទដែលកាន់កាប់តំបន់អេកូឡូស៊ីដូចគ្នា (រូបភាព 9.3 a, b) និងប្រភេទសត្វដែលកាន់កាប់តំបន់អេកូឡូស៊ីខុសៗគ្នា (រូបភាព 9.3 គ)។
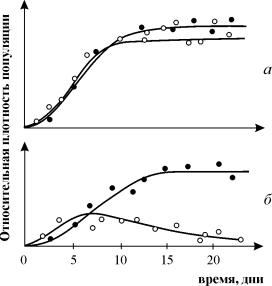
អង្ករ។ ៩.៣. ក- ខ្សែកោងកំណើនប្រជាជននៃប្រភេទពីរប៉ារ៉ាមេន នៅក្នុងដំណាំតែមួយប្រភេទ។ រង្វង់ខ្មៅ - P Aurelia, រង្វង់ពណ៌ស - P. Caudatum
ខ- ខ្សែកោងកំណើននៃ P Aurelia និង P . Caudatum នៅក្នុងវប្បធម៌ចម្រុះ។
ដោយ Gause ឆ្នាំ 1934
គំរូនៃការប្រកួតប្រជែង (9.2) មានគុណវិបត្តិ ជាពិសេសវាកើតឡើងថាការរួមរស់នៃប្រភេទសត្វពីរគឺអាចធ្វើទៅបានលុះត្រាតែចំនួនរបស់វាត្រូវបានកំណត់ដោយកត្តាផ្សេងៗគ្នា ប៉ុន្តែគំរូមិនបង្ហាញពីទំហំភាពខុសគ្នាដែលត្រូវតែមានដើម្បីធានាការរួមរស់បានយូរនោះទេ។ . ក្នុងពេលជាមួយគ្នានេះ វាត្រូវបានគេដឹងថាសម្រាប់ការរួមរស់ជាមួយគ្នាយូរអង្វែងនៅក្នុងបរិយាកាសផ្លាស់ប្តូរ ភាពខុសគ្នាឈានដល់ទំហំជាក់លាក់មួយគឺចាំបាច់។ ការណែនាំធាតុ stochastic ទៅក្នុងគំរូ (ឧទាហរណ៍ ការណែនាំមុខងារប្រើប្រាស់ធនធាន) អនុញ្ញាតឱ្យយើងធ្វើការស៊ើបអង្កេតលើបញ្ហាទាំងនេះជាបរិមាណ។
ប្រព័ន្ធ PREDATOR + VICTIM
 (9.7)
(9.7)
នៅទីនេះផ្ទុយទៅនឹង (9.2) សញ្ញា ខ 12 និង ខ 21 គឺខុសគ្នា។ ដូចនៅក្នុងករណីនៃការប្រកួតប្រជែង, ប្រភពដើម
![]() (9.8)
(9.8)
គឺជាចំណុចពិសេសនៃប្រភេទថ្នាំងមិនស្ថិតស្ថេរ។ ស្ថានភាពស្ថិរភាពបីផ្សេងទៀត៖
![]() ,(9.9)
,(9.9)
![]() (9.10)
(9.10)
![]() (9.11)
(9.11)
ដូច្នេះវាអាចទៅរួចសម្រាប់តែសត្វព្រៃដែលអាចរស់បាន (9.10) មានតែសត្វមំសាសី (9.9) (ប្រសិនបើវាមានប្រភពអាហារផ្សេងទៀត) និងការរស់នៅជាមួយគ្នានៃប្រភេទទាំងពីរ (9.11) ។ យើងបានពិភាក្សាអំពីជម្រើសចុងក្រោយរួចហើយនៅក្នុងមេរៀនទី 5 ។ ប្រភេទដែលអាចកើតមាននៃរូបភាពដំណាក់កាលសម្រាប់ប្រព័ន្ធ predator-prey ត្រូវបានបង្ហាញនៅក្នុងរូបភព។ ៩.៤.
Isoclins នៃតង់ហ្សង់ផ្ដេកគឺជាបន្ទាត់ត្រង់
x 2 = – ខ 21 X 1 /គ 2 + ក១/គ ២, X 2 = 0,
និង isoclines នៃតង់សង់បញ្ឈរ- ត្រង់
x 2 = – គ 1 X 1 /ខ 12 + ក 2 /ខ 12 , X 1 = 0.
ចំនុចស្ថានីស្ថិតនៅចំនុចប្រសព្វនៃ isoclines តង់ហ្សង់បញ្ឈរ និងផ្ដេក។

ពីរូបភព។ ៩.៤ ខាងក្រោមនេះអាចមើលឃើញ។ ប្រព័ន្ធ Predator-prey (9.7) អាចមានទីតាំងលំនឹងថេរ ដែលក្នុងនោះ o Rum ប្រជាជនជនរងគ្រោះបានស្លាប់ទាំងស្រុង ( ) ហើយនៅសល់តែសត្វមំសាសីប៉ុណ្ណោះ (រយៈពេល 2 នៅក្នុងរូបភព។ ៩.៤ ក). ជាក់ស្តែង ស្ថានភាពបែបនេះអាចដឹងបានលុះត្រាតែបន្ថែមលើប្រភេទនៃជនរងគ្រោះនៅក្នុងសំណួរ។ X 1 predator X២- មានប្រភពថាមពលបន្ថែម។ ការពិតនេះត្រូវបានឆ្លុះបញ្ចាំងនៅក្នុងគំរូដោយពាក្យវិជ្ជមាននៅជ្រុងខាងស្តាំនៃសមីការសម្រាប់ x2 ។ ចំណុចពិសេស(1) និង (3) (រូបភាព 9.4 ក) មិនស្ថិតស្ថេរ។ លទ្ធភាពទីពីរ – ស្ថានភាពស្ថិរភាពដែលចំនួនសត្វមំសាសីបានស្លាប់ទាំងស្រុង ហើយនៅសល់តែសត្វព្រៃប៉ុណ្ណោះ។ – ចំណុចមានស្ថេរភាព(3) (រូបភាព 9.4 6 ). មានចំណុចពិសេសមួយនៅទីនេះ (1) – ថ្នាំងមិនស្ថិតស្ថេរផងដែរ។
ទីបំផុតលទ្ធភាពទីបី – ការរួមរស់ជាមួយគ្នាប្រកបដោយនិរន្តរភាពនៃចំនួនប្រជាជននៃសត្វមំសាសី និងសត្វព្រៃ (រូបភាពទី. 9.4 វ) លេខស្ថានីដែលត្រូវបានបង្ហាញដោយរូបមន្ត (9.11).
ដូចនៅក្នុងករណីនៃចំនួនប្រជាជនមួយ (សូមមើល មេរៀនទី 3) សម្រាប់គំរូ (9.7) វាអាចទៅរួចក្នុងការអភិវឌ្ឍគំរូ stochastic ប៉ុន្តែវាមិនអាចដោះស្រាយបានច្បាស់លាស់ទេ។ ដូច្នេះ យើងនឹងដាក់កម្រិតខ្លួនយើងចំពោះការពិចារណាទូទៅ។ ជាឧទាហរណ៍ ចូរយើងសន្មត់ថា ចំណុចលំនឹងស្ថិតនៅចម្ងាយជាក់លាក់មួយពីអ័ក្សនីមួយៗ។ បន្ទាប់មកសម្រាប់គន្លងដំណាក់កាលដែលតម្លៃx 1 , x 2 នៅតែមានទំហំធំល្មម គំរូកំណត់នឹងពេញចិត្តណាស់។ ប៉ុន្តែប្រសិនបើនៅចំណុចមួយចំនួននៅក្នុងគន្លងដំណាក់កាលអថេរណាមួយមិនមានទំហំធំទេនោះ ការប្រែប្រួលចៃដន្យអាចក្លាយជាសំខាន់។ ពួកវានាំទៅរកការពិតដែលថាចំណុចតំណាងផ្លាស់ទីទៅអ័ក្សមួយក្នុងចំណោមអ័ក្សដែលមានន័យថាការផុតពូជនៃប្រភេទសត្វដែលត្រូវគ្នា។
ដូច្នេះគំរូ stochastic ប្រែទៅជាមិនស្ថិតស្ថេរចាប់តាំងពី "រសាត់" stochastic ឆាប់ឬក្រោយមកនាំទៅដល់ការផុតពូជនៃប្រភេទសត្វមួយ។ នៅក្នុងគំរូប្រភេទនេះ សត្វមំសាសីនៅទីបំផុតបានផុតពូជ ទោះដោយចៃដន្យ ឬដោយសារចំនួនសត្វព្រៃរបស់វាត្រូវបានកំចាត់មុនគេ។ គំរូ stochastic នៃប្រព័ន្ធ predator-prey ពន្យល់ពីការពិសោធន៍របស់ Gause បានយ៉ាងល្អ (Gause, 1934), ដែលក្នុងនោះ ciliates Paramettum candatumបានបម្រើជាជនរងគ្រោះសម្រាប់ ciliate មួយផ្សេងទៀត ឌីឌីនីញ៉ូម ណាសាតម – មំសាសី។ រំពឹងយោងទៅតាមសមីការកំណត់ (9.7) លេខលំនឹងនៅក្នុងការពិសោធន៍ទាំងនេះមានប្រមាណតែ 5 នាក់ប៉ុណ្ណោះនៃប្រភេទនីមួយៗ ដូច្នេះវាមិនគួរឱ្យភ្ញាក់ផ្អើលទេដែលនៅក្នុងការពិសោធន៍ម្តងហើយម្តងទៀត ទាំងសត្វមំសាសី ឬសត្វព្រៃ (ហើយបន្ទាប់ពីពួកវា មំសាសី) បានស្លាប់យ៉ាងឆាប់រហ័ស លទ្ធផលនៃការពិសោធន៍ត្រូវបានបង្ហាញ នៅក្នុងរូបភព។ ៩.៥.

អង្ករ. 9.5. កម្ពស់ ប៉ារ៉ាមីញ៉ូម caudatum និង ciliates predatory ដាឌីនីញ៉ូម. ពី ៖ Gause G.F. ការតស៊ូដើម្បីអត្ថិភាព។ បាល់ទីម័រ, 1934
ដូច្នេះការវិភាគនៃគំរូ Volterra នៃអន្តរកម្មនៃប្រភេទសត្វបង្ហាញថា ទោះបីជាមានភាពខុសគ្នានៃប្រភេទនៃឥរិយាបទនៃប្រព័ន្ធបែបនេះក៏ដោយ ក៏មិនអាចមានភាពប្រែប្រួលនៃចំនួននៅក្នុងគំរូនៃប្រភេទសត្វដែលប្រកួតប្រជែងគ្នាបានឡើយ។ ទោះជាយ៉ាងណាក៏ដោយ លំយោលបែបនេះត្រូវបានគេសង្កេតឃើញនៅក្នុងធម្មជាតិ និងនៅក្នុងការពិសោធន៍។ តម្រូវការសម្រាប់ការពន្យល់ទ្រឹស្តីរបស់ពួកគេគឺជាហេតុផលមួយសម្រាប់បង្កើតការពិពណ៌នាគំរូក្នុងទម្រង់ទូទៅជាង។
គំរូទូទៅនៃអន្តរកម្មនៃប្រភេទពីរ
គំរូមួយចំនួនធំត្រូវបានស្នើឡើងដើម្បីពិពណ៌នាអំពីអន្តរកម្មនៃប្រភេទសត្វ ដែលផ្នែកខាងស្តាំនៃសមីការដែលជាមុខងារនៃចំនួនប្រជាជនអន្តរកម្ម។ បញ្ហានៃការបង្កើតលក្ខណៈវិនិច្ឆ័យទូទៅដើម្បីបង្កើតមុខងារប្រភេទណាដែលអាចពិពណ៌នាអំពីឥរិយាបថនៃទំហំប្រជាជនបណ្តោះអាសន្ន រួមទាំងការប្រែប្រួលស្ថិរភាពត្រូវបានដោះស្រាយ។ ម៉ូដែលដ៏ល្បីល្បាញបំផុតទាំងនេះជាកម្មសិទ្ធិរបស់ Kolmogorov (1935, អត្ថបទកែប្រែ - 1972) និង Rosenzweig (1963) ។
 (9.12)
(9.12)
គំរូរួមបញ្ចូលការសន្មត់ដូចខាងក្រោមៈ
1) Predators មិនទាក់ទងគ្នាទៅវិញទៅមក, i.e. អត្រាបន្តពូជរបស់សត្វខ្លា k២ និងចំនួនជនរងគ្រោះ អិល exterminated ក្នុងមួយឯកតាពេលវេលាដោយ predator មួយមិនអាស្រ័យលើ y.
2) ការកើនឡើងនៃចំនួនសត្វព្រៃនៅក្នុងវត្តមានរបស់សត្វមំសាសីគឺស្មើនឹងការកើនឡើងនៃអវត្ដមាននៃមំសាសី ដកចំនួនសត្វឈ្មោលដែលត្រូវបានសម្លាប់ដោយមំសាសី។ មុខងារ k 1 (x), k 2 (x), អិល(x) ត្រូវបានបន្ត និងកំណត់នៅលើអ័ក្សពាក់កណ្តាលវិជ្ជមាន x, y³ 0.
3) ឃ 1 /dx< 0. នេះមានន័យថា អត្រានៃការបន្តពូជរបស់សត្វព្រៃ ក្នុងពេលគ្មានសត្វមំសាសី មានការថយចុះជាឯកតោភាគី ជាមួយនឹងការកើនឡើងនៃចំនួនសត្វព្រៃ ដែលឆ្លុះបញ្ចាំងពីភាពអាចរកបានតិចតួចនៃអាហារ និងធនធានផ្សេងទៀត។
4) ឃ 2 /dx> 0, ក 2 (0) < 0 < k 2 (¥ ). ជាមួយនឹងការកើនឡើងនៃចំនួនសត្វឈ្មោល មេគុណបន្តពូជរបស់សត្វមំសាសីមានការថយចុះដោយឯកឯងជាមួយនឹងការកើនឡើងនៃចំនួនសត្វព្រៃ ដោយផ្លាស់ប្តូរពី តម្លៃអវិជ្ជមាន, (នៅពេលដែលគ្មានអ្វីបរិភោគ) ទៅជាវិជ្ជមាន។
5) ចំនួនសត្វព្រៃដែលត្រូវបានបំផ្លាញដោយសត្វមំសាសីម្នាក់ក្នុងមួយឯកតានៃពេលវេលា អិល(x)> 0 នៅ ន > 0; អិល(0)=0.
ប្រភេទដែលអាចកើតមាននៃរូបភាពដំណាក់កាលនៃប្រព័ន្ធ (9.12) ត្រូវបានបង្ហាញនៅក្នុងរូបភព។ ៩.៦៖

អង្ករ។ ៩.៦.រូបថតដំណាក់កាលនៃប្រព័ន្ធ Kolmogorov (9.12) ដែលពិពណ៌នាអំពីអន្តរកម្មនៃប្រភេទពីរនៅសមាមាត្រផ្សេងគ្នានៃប៉ារ៉ាម៉ែត្រ។ ការពន្យល់នៅក្នុងអត្ថបទ។
ដំណោះស្រាយស្ថានី (មានពីរឬបី) មានកូអរដោនេដូចខាងក្រោមៈ
(1). ` x=0;` y=0.
ប្រភពដើមនៃកូអរដោណេសម្រាប់តម្លៃប៉ារ៉ាម៉ែត្រណាមួយគឺ ក្រវ៉ាត់ (រូបភាព 9.6 a-d) ។
(2). ` x=A,` y=0។(9.13)
កកំណត់ពីសមីការ៖
k 1 (ក)=0.
ស្ថានី ដំណោះស្រាយ (៩.១៣) គឺជាក្រវ៉ាត់មួយប្រសិនបើ ខ< ក (រូបភាព 9.6 ក, ខ, ជី), ខ កំណត់ពីសមីការ
k 2 (ខ)=0
ចំណុច (9.13) ត្រូវបានដាក់ក្នុង quadrant វិជ្ជមាន ប្រសិនបើ B> ក . នេះគឺជាថ្នាំងដែលមានស្ថេរភាព .
ករណីចុងក្រោយ ដែលត្រូវនឹងការស្លាប់របស់សត្វមំសាសី និងការរស់រានមានជីវិតរបស់សត្វព្រៃ ត្រូវបានបង្ហាញនៅក្នុងរូបភព។ 9.6 វ.
(3). ` x=B,` y=C(9.14)
តម្លៃ C ត្រូវបានកំណត់ដោយសមីការ៖
ចំណុច (៩.១៤) - ការផ្តោតអារម្មណ៍ (រូបភាព ៩.៦ ក) ឬថ្នាំង (Fig.9.6 ជី) ស្ថេរភាពដែលអាស្រ័យលើសញ្ញានៃបរិមាណស
ស 2 = – k 1 (ខ) - គ 1 (ខ)B+L(ខ)គ.
ប្រសិនបើ ស>0, ចំណុចមួយមានស្ថេរភាពប្រសិនបើស<0 ‑ точка неустойчива, и вокруг нее могут существовать предельные циклы (рис. 9.6 ខ)
នៅក្នុងអក្សរសិល្ប៍បរទេស គំរូស្រដៀងគ្នាដែលស្នើឡើងដោយ Rosenzweig និង MacArthur (1963) ត្រូវបានគេចាត់ទុកថាញឹកញាប់ជាង៖
 (9.15)
(9.15)
កន្លែងណា f(x) - អត្រានៃការផ្លាស់ប្តូរចំនួនជនរងគ្រោះ xអវត្ដមាននៃសត្វមំសាសី F( x, y) - អាំងតង់ស៊ីតេនៃការឈ្លានពាន k- មេគុណកំណត់លក្ខណៈប្រសិទ្ធភាពនៃការកែច្នៃជីវម៉ាសទៅជាជីវម៉ាសសត្វ អ៊ី- ការស្លាប់របស់សត្វមំសាសី។
គំរូ (9.15) កាត់បន្ថយទៅជាករណីពិសេសនៃគំរូ Kolmogorov (9.12) ក្រោមការសន្មត់ដូចខាងក្រោម:
1) ចំនួនសត្វមំសាសីត្រូវបានកំណត់ត្រឹមចំនួនសត្វព្រៃប៉ុណ្ណោះ
2) ល្បឿនដែលសត្វមំសាសីបានស៊ីចំណីគឺអាស្រ័យតែលើដង់ស៊ីតេនៃចំនួនសត្វព្រៃប៉ុណ្ណោះ និងមិនអាស្រ័យលើដង់ស៊ីតេនៃចំនួនសត្វមំសាសីនោះទេ។
បន្ទាប់មកសមីការ (៩.១៥) យកទម្រង់។

នៅពេលពិពណ៌នាអំពីអន្តរកម្មនៃប្រភេទសត្វពិត ជ្រុងខាងស្តាំនៃសមីការត្រូវបានបញ្ជាក់ដោយអនុលោមតាមគំនិតអំពីការពិតជីវសាស្រ្ត។ ចូរយើងពិចារណាអំពីម៉ូដែលដ៏ពេញនិយមបំផុតមួយនៃប្រភេទនេះ។
គំរូនៃអន្តរកម្មរវាងសត្វល្អិតពីរប្រភេទ (MacArthur, 1971)
គំរូដែលយើងនឹងពិចារណាខាងក្រោម ត្រូវបានប្រើដើម្បីដោះស្រាយបញ្ហាជាក់ស្តែងនៃការគ្រប់គ្រងសត្វល្អិតដែលបង្កគ្រោះថ្នាក់ដោយការក្រៀវឈ្មោលនៃប្រភេទសត្វមួយ។ ដោយផ្អែកលើលក្ខណៈជីវសាស្រ្តនៃអន្តរកម្មប្រភេទសត្វ គំរូខាងក្រោមត្រូវបានសរសេរ
 (9.16)
(9.16)
នៅទីនេះ x, y- ជីវម៉ាសនៃសត្វល្អិតពីរប្រភេទ។ អន្តរកម្ម trophic នៃប្រភេទដែលបានពិពណ៌នានៅក្នុងគំរូនេះគឺស្មុគស្មាញណាស់។ វាកំណត់ទម្រង់នៃពហុនាមនៅខាងស្តាំដៃនៃសមីការ។
សូមក្រឡេកមើលផ្នែកខាងស្តាំនៃសមីការទីមួយ។ ប្រភេទសត្វល្អិត Xបរិភោគដង្កូវនៃប្រភេទសត្វ នៅ(សមាជិក +k 3 y)ប៉ុន្តែមនុស្សពេញវ័យនៃប្រភេទសត្វ នៅបរិភោគដង្កូវនៃប្រភេទសត្វ Xភាពសម្បូរបែបនៃប្រភេទសត្វខ្ពស់។ Xឬ នៅឬទាំងពីរប្រភេទ (សមាជិក – គ 4 xy, – y ២). នៅតូច Xការស្លាប់នៃប្រភេទសត្វ Xខ្ពស់ជាងការកើនឡើងធម្មជាតិរបស់វា (១ -k 1 +k 2 x–x ២ < 0 នៅតូច X).នៅក្នុងសមីការទីពីរពាក្យ k 5 ឆ្លុះបញ្ចាំងពីការលូតលាស់ធម្មជាតិនៃប្រភេទសត្វ y; -k 6 y –ការអត់ធ្មត់ដោយខ្លួនឯងនៃប្រភេទនេះ,-k 7 x- បរិភោគដង្កូវនៃប្រភេទសត្វ នៅប្រភេទសត្វល្អិត x, ក 8 xy – ការកើនឡើងនៃប្រភេទជីវម៉ាស នៅដោយសារតែការទទួលទានដោយសត្វល្អិតពេញវ័យនៃប្រភេទសត្វ នៅ larvae នៃប្រភេទ X.
នៅក្នុងរូបភព។ ៩.៧ វដ្តដែនកំណត់ត្រូវបានបង្ហាញ ដែលជាគន្លងនៃដំណោះស្រាយតាមកាលកំណត់ដែលមានស្ថេរភាពនៃប្រព័ន្ធ (9.16).

ដំណោះស្រាយចំពោះសំណួរអំពីរបៀបធានាឱ្យមានការរួមរស់ជាមួយគ្នានៃចំនួនប្រជាជនជាមួយនឹងបរិស្ថានជីវសាស្រ្តរបស់វា មិនអាចទទួលបានដោយមិនគិតគូរពីភាពជាក់លាក់នៃប្រព័ន្ធជីវសាស្ត្រជាក់លាក់មួយ និងការវិភាគនៃទំនាក់ទំនងអន្តរកម្មរបស់វាទាំងអស់។ ទន្ទឹមនឹងនេះ ការសិក្សាអំពីគំរូគណិតវិទ្យាផ្លូវការអនុញ្ញាតឱ្យយើងឆ្លើយសំណួរទូទៅមួយចំនួន។ វាអាចត្រូវបានអះអាងថាសម្រាប់ម៉ូដែលដូចជា (9.12) ការពិតនៃភាពឆបគ្នាឬភាពមិនស៊ីគ្នានៃចំនួនប្រជាជនមិនអាស្រ័យលើទំហំដំបូងរបស់ពួកគេទេប៉ុន្តែត្រូវបានកំណត់ដោយធម្មជាតិនៃអន្តរកម្មនៃប្រភេទសត្វប៉ុណ្ណោះ។ គំរូជួយឆ្លើយសំណួរ៖ របៀបជះឥទ្ធិពលលើ biocenosis និងគ្រប់គ្រងវា ដើម្បីបំផ្លាញប្រភេទសត្វដែលបង្កគ្រោះថ្នាក់យ៉ាងឆាប់រហ័ស។
ការគ្រប់គ្រងអាចត្រូវបានកាត់បន្ថយមកត្រឹមរយៈពេលខ្លី ការផ្លាស់ប្តូរភ្លាមៗនៃតម្លៃប្រជាជន Xនិង យូវិធីសាស្រ្តនេះត្រូវគ្នាទៅនឹងវិធីសាស្ត្រគ្រប់គ្រង ដូចជាការបំផ្លិចបំផ្លាញតែម្តងនៃចំនួនប្រជាជនមួយ ឬទាំងពីរដោយមធ្យោបាយគីមី។ តាមសេចក្តីថ្លែងការណ៍ដែលបានបង្កើតខាងលើ វាច្បាស់ណាស់ថាសម្រាប់ប្រជាជនដែលត្រូវគ្នា វិធីសាស្ត្រនៃការគ្រប់គ្រងនេះនឹងគ្មានប្រសិទ្ធភាពទេ ព្រោះយូរៗទៅប្រព័ន្ធនឹងឈានដល់របបស្ថានីម្តងទៀត។
វិធីមួយទៀតគឺផ្លាស់ប្តូរប្រភេទនៃមុខងារអន្តរកម្មរវាងទិដ្ឋភាពឧទាហរណ៍នៅពេលផ្លាស់ប្តូរតម្លៃនៃប៉ារ៉ាម៉ែត្រប្រព័ន្ធ។ វាគឺជាវិធីសាស្រ្តប៉ារ៉ាម៉ែត្រនេះដែលវិធីសាស្ត្រគ្រប់គ្រងជីវសាស្រ្តត្រូវគ្នា។ ដូច្នេះហើយ នៅពេលដែលមេអំបៅត្រូវបានណែនាំ អត្រានៃកំណើនប្រជាជនធម្មជាតិមានការថយចុះ។ ប្រសិនបើក្នុងពេលជាមួយគ្នានេះ យើងទទួលបានប្រភេទបញ្ឈរនៃដំណាក់កាលផ្សេងគ្នា ដែលកន្លែងមួយដែលមានតែស្ថានភាពថេរដែលមានលេខសត្វល្អិតសូន្យ ការគ្រប់គ្រងនឹងនាំទៅរកលទ្ធផលដែលចង់បាន។ – ការបំផ្លាញចំនួនប្រជាជននៃប្រភេទសត្វដែលបង្កគ្រោះថ្នាក់។ វាគួរឱ្យចាប់អារម្មណ៍ក្នុងការកត់សម្គាល់ថាជួនកាលវាត្រូវបានគេណែនាំអោយអនុវត្តផលប៉ះពាល់មិនមែនចំពោះសត្វល្អិតខ្លួនឯងទេតែចំពោះដៃគូរបស់វា។ ជាទូទៅ គេមិនអាចនិយាយបានថា វិធីសាស្ត្រមួយណាមានប្រសិទ្ធភាពជាង។ វាអាស្រ័យលើការគ្រប់គ្រងដែលមាន និងនៅលើទម្រង់ច្បាស់លាស់នៃមុខងារដែលពិពណ៌នាអំពីអន្តរកម្មនៃចំនួនប្រជាជន។
ម៉ូដែល A.D. Bazykin
ការវិភាគទ្រឹស្តីនៃគំរូនៃអន្តរកម្មនៃប្រភេទសត្វត្រូវបានអនុវត្តយ៉ាងទូលំទូលាយបំផុតនៅក្នុងសៀវភៅរបស់ AD Bazykin "Biophysics of Interacting Populations" (M., Nauka, 1985) ។
សូមពិចារណាគំរូសត្វឈ្មោលមួយដែលបានសិក្សាក្នុងសៀវភៅនេះ។
 (9.17)
(9.17)
ប្រព័ន្ធ (9.17) គឺជាការធ្វើឱ្យទូទៅនៃគំរូ Volterra predator-prey សាមញ្ញបំផុត (5.17) ដោយគិតគូរពីឥទ្ធិពលនៃការតិត្ថិភាពនៃសត្វមំសាសី។ គំរូ (5.17) សន្មត់ថា អាំងតង់ស៊ីតេនៃការស៊ីស្មៅកើនឡើងស្របគ្នាជាមួយនឹងការកើនឡើងនៃដង់ស៊ីតេសត្វព្រៃ ដែលមិនស៊ីគ្នានឹងការពិតនៅដង់ស៊ីតេសត្វព្រៃខ្ពស់។ មុខងារផ្សេងគ្នាអាចត្រូវបានជ្រើសរើសដើម្បីពិពណ៌នាអំពីភាពអាស្រ័យនៃរបបអាហាររបស់សត្វមំសាសី លើដង់ស៊ីតេសត្វព្រៃ។ វាសំខាន់បំផុតដែលមុខងារដែលបានជ្រើសរើសជាមួយនឹងការលូតលាស់ xទំនោរ asymptotically ទៅតម្លៃថេរ។ គំរូ (9.6) បានប្រើការពឹងផ្អែកនៃភស្តុភារ។ នៅក្នុងគំរូរបស់ Bazykin អ៊ីពែបូឡាត្រូវបានជ្រើសរើសជាមុខងារបែបនេះ x/(1+px) ចូរយើងចងចាំថានេះគឺជាទម្រង់នៃរូបមន្ត Monod ដែលពិពណ៌នាអំពីភាពអាស្រ័យនៃអត្រាកំណើននៃ microorganisms លើកំហាប់នៃស្រទាប់ខាងក្រោម។ នៅទីនេះ សត្វពាហនៈដើរតួជាស្រទាប់ខាងក្រោម ហើយមំសាសីដើរតួជាអតិសុខុមប្រាណ។ .
ប្រព័ន្ធ (9.17) អាស្រ័យលើប៉ារ៉ាម៉ែត្រប្រាំពីរ។ ចំនួនប៉ារ៉ាម៉ែត្រអាចត្រូវបានកាត់បន្ថយដោយការជំនួសអថេរ៖
x® (ក/ឃ)x; y ® (ក/ឃ)/y;
t® (១/ ក)t; g (9.18)
និងអាស្រ័យលើប៉ារ៉ាម៉ែត្រចំនួនបួន។
សម្រាប់ការសិក្សាគុណភាពពេញលេញ វាចាំបាច់ក្នុងការបែងចែកចន្លោះប៉ារ៉ាម៉ែត្របួនវិមាត្រទៅជាតំបន់ដែលមានប្រភេទផ្សេងគ្នានៃឥរិយាបទថាមវន្ត ពោលគឺឧ។ បង្កើតប៉ារ៉ាម៉ែត្រ ឬរចនាសម្ព័ន្ធបញ្ឈរនៃប្រព័ន្ធ។
បន្ទាប់មក វាចាំបាច់ក្នុងការសង់រូបតំណាក់កាលសម្រាប់តំបន់នីមួយៗនៃរូបបញ្ឈរប៉ារ៉ាមេត ហើយពិពណ៌នាអំពីភាពច្របូកច្របល់ដែលកើតឡើងជាមួយនឹងរូបតំណាក់កាលនៅព្រំដែននៃតំបន់ផ្សេងៗគ្នានៃរូបប៉ារ៉ាមេត។
ការសាងសង់រូបបញ្ឈរប៉ារ៉ាមេតពេញលេញត្រូវបានអនុវត្តជាទម្រង់នៃ "ចំណិត" (ការព្យាករណ៍) នៃរូបបញ្ឈរប៉ារ៉ាម៉ែត្រវិមាត្រទាបជាមួយនឹងតម្លៃថេរនៃប៉ារ៉ាម៉ែត្រមួយចំនួន។
ប៉ារ៉ាម៉ែត្របញ្ឈរនៃប្រព័ន្ធ (9.18) សម្រាប់ថេរ gនិងតូច អ៊ីបង្ហាញក្នុងរូប ៩.៨។ រូបបញ្ឈរមាន 10 តំបន់ដែលមានប្រភេទផ្សេងគ្នានៃឥរិយាបទនៃគន្លងដំណាក់កាល។

អង្ករ។ ៩.៨.ប៉ារ៉ាម៉ែត្របញ្ឈរនៃប្រព័ន្ធ (9.18) សម្រាប់ថេរg
និងតូច អ៊ី
ឥរិយាបថនៃប្រព័ន្ធនៅសមាមាត្រផ្សេងគ្នានៃប៉ារ៉ាម៉ែត្រអាចមានភាពខុសគ្នាខ្លាំង (រូបភាព 9.9) ។ ប្រព័ន្ធអនុញ្ញាតឱ្យ៖
1) លំនឹងស្ថិរភាពមួយ (តំបន់ 1 និង 5);
2) វដ្តដែនកំណត់ស្ថិរភាពមួយ (តំបន់ 3 និង 8);
3) លំនឹងស្ថេរភាពពីរ (តំបន់ 2)
4) វដ្តដែនកំណត់មានស្ថេរភាព និងលំនឹងមិនស្ថិតស្ថេរនៅខាងក្នុងវា (តំបន់ 6, 7, 9, 10)
5) វដ្តដែនកំណត់មានស្ថេរភាព និងលំនឹងស្ថិរភាពនៅខាងក្រៅវា (តំបន់ទី 4) ។
នៅក្នុងតំបន់ប៉ារ៉ាម៉ែត្រ 7, 9, 10 តំបន់នៃការទាក់ទាញនៃលំនឹងត្រូវបានកំណត់ដោយវដ្តដែនកំណត់មិនស្ថិតស្ថេរដែលស្ថិតនៅក្នុងស្ថេរភាពមួយ។ រចនាសម្ព័ន្ធគួរឱ្យចាប់អារម្មណ៍បំផុតគឺបញ្ឈរដំណាក់កាលដែលត្រូវគ្នានឹងផ្ទៃ 6 នៅក្នុងបញ្ឈរប៉ារ៉ាម៉ែត្រ។ វាត្រូវបានបង្ហាញលម្អិតនៅក្នុងរូបភព។ ៩.១០.
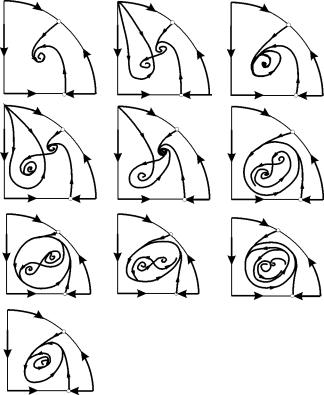
តំបន់ទាក់ទាញនៃលំនឹង B 2 (ស្រមោល) គឺជា "ខ្យង" បង្វិលពីការផ្តោតអារម្មណ៍មិនស្ថិតស្ថេរ B 1 ។ ប្រសិនបើគេដឹងថានៅដំណាក់កាលដំបូងនៃប្រព័ន្ធគឺនៅជិត B 1 នោះគេអាចវិនិច្ឆ័យថាតើគន្លងដែលត្រូវគ្នានឹងឈានដល់លំនឹង B 2 ឬវដ្តដែនកំណត់ដែលមានស្ថេរភាពជុំវិញចំនុចលំនឹងទាំងបី C (saddle ), B 1 និង B 2 ផ្អែកលើការពិចារណាអំពីប្រូបាប៊ីលីតេ។

រូប ៩.១០.ដំណាក់កាលបញ្ឈរនៃប្រព័ន្ធ 9.18 សម្រាប់តំបន់ប៉ារ៉ាម៉ែត្រ 6. តំបន់ទាក់ទាញ B 2 ត្រូវបានដាក់ស្រមោល
នៅក្នុងរូបថតប៉ារ៉ាម៉ែត្រ(៩.៧) មាន ២២ ព្រំដែន bifurcation ផ្សេងគ្នាដែលបង្កើត 7 ប្រភេទផ្សេងគ្នានៃ bifurcations ។ ការសិក្សារបស់ពួកគេអនុញ្ញាតឱ្យយើងកំណត់ប្រភេទនៃឥរិយាបទប្រព័ន្ធដែលអាចកើតមាននៅពេលដែលប៉ារ៉ាម៉ែត្ររបស់វាផ្លាស់ប្តូរ។ ឧទាហរណ៍នៅពេលផ្លាស់ទីពីតំបន់ 1 ដល់តំបន់ 3 កំណើតនៃវដ្តដែនកំណត់តូចមួយកើតឡើង ឬកំណើតទន់នៃការយោលដោយខ្លួនឯងជុំវិញលំនឹងតែមួយ INកំណើតទន់ស្រដៀងគ្នានៃការយោលដោយខ្លួនឯង ប៉ុន្តែនៅជុំវិញលំនឹងមួយ ពោលគឺ ខ 1 , កើតឡើងនៅពេលឆ្លងកាត់ព្រំដែននៃតំបន់ 2 និង 4 ។ នៅពេលចាកចេញពីតំបន់៤ ទៅតំបន់ ៥ រង្វង់ដែនកំណត់មានស្ថេរភាពជុំវិញចំណុចមួយ។ខ 1 "ផ្ទុះ" នៅលើរង្វិលជុំនៃសញ្ញាបំបែកហើយចំណុចទាក់ទាញតែមួយគត់នៅតែមានលំនឹង ខ 2 ល។
ការចាប់អារម្មណ៍ជាពិសេសសម្រាប់ការអនុវត្តគឺជាការពិតណាស់ ការអភិវឌ្ឍន៍លក្ខណៈវិនិច្ឆ័យសម្រាប់ភាពជិតនៃប្រព័ន្ធទៅនឹងព្រំដែន bifurcation ។ ជាការពិត អ្នកជីវវិទូដឹងយ៉ាងច្បាស់អំពីទ្រព្យសម្បត្តិ "រារាំង" ឬ "ភាពបត់បែន" នៃប្រព័ន្ធអេកូឡូស៊ីធម្មជាតិ។ ពាក្យទាំងនេះជាធម្មតាសំដៅទៅលើសមត្ថភាពនៃប្រព័ន្ធមួយក្នុងការស្រូបយកឥទ្ធិពលខាងក្រៅ។ ដរាបណាអាំងតង់ស៊ីតេនៃឥទ្ធិពលខាងក្រៅមិនលើសពីតម្លៃសំខាន់ជាក់លាក់មួយ ឥរិយាបថនៃប្រព័ន្ធមិនឆ្លងកាត់ការផ្លាស់ប្តូរគុណភាពទេ។ នៅលើយន្តហោះដំណាក់កាល នេះត្រូវគ្នាទៅនឹងការវិលត្រឡប់នៃប្រព័ន្ធទៅស្ថានភាពលំនឹងស្ថិរភាព ឬវដ្តដែនកំណត់ដែលមានស្ថេរភាព ប៉ារ៉ាម៉ែត្រដែលមិនខុសគ្នាច្រើនពីប្រព័ន្ធដើម។ នៅពេលដែលអាំងតង់ស៊ីតេនៃផលប៉ះពាល់លើសពីកម្រិតដែលអាចអនុញ្ញាតបាននោះ ប្រព័ន្ធ "ខូច" ហើយចូលទៅក្នុងរបៀបនៃឥរិយាបទថាមវន្តដែលមានលក្ខណៈខុសៗគ្នា ឧទាហរណ៍ វាគ្រាន់តែងាប់។ បាតុភូតនេះត្រូវគ្នាទៅនឹងការផ្លាស់ប្តូរ bifurcation ។
ប្រភេទនៃការផ្លាស់ប្តូរ bifurcation នីមួយៗមានលក្ខណៈពិសេសផ្ទាល់ខ្លួនរបស់វា ដែលធ្វើឱ្យវាអាចធ្វើទៅបានដើម្បីវិនិច្ឆ័យគ្រោះថ្នាក់នៃការផ្លាស់ប្តូរបែបនេះសម្រាប់ប្រព័ន្ធអេកូ។ នេះគឺជាលក្ខណៈវិនិច្ឆ័យទូទៅមួយចំនួនដែលបង្ហាញពីភាពជិតនៃព្រំដែនដែលមានគ្រោះថ្នាក់។ ដូចនៅក្នុងករណីនៃប្រភេទសត្វមួយ ប្រសិនបើនៅពេលដែលចំនួននៃប្រភេទសត្វមួយមានការថយចុះ ប្រព័ន្ធនេះ "ជាប់គាំង" នៅជិតចំនុចកួរមិនស្ថិតស្ថេរ ដែលត្រូវបានបញ្ជាក់នៅក្នុងការស្ដារឡើងវិញយឺតបំផុតនៃចំនួនទៅតម្លៃដំបូង បន្ទាប់មក ប្រព័ន្ធគឺនៅជិតព្រំដែនសំខាន់។ សូចនាករនៃគ្រោះថ្នាក់ក៏ជាការផ្លាស់ប្តូររូបរាងប្រែប្រួលនៃចំនួនសត្វមំសាសី និងសត្វព្រៃផងដែរ។ ប្រសិនបើលំយោលដែលនៅជិតនឹងអាម៉ូនិកក្លាយជាការបន្ធូរបន្ថយ ហើយទំហំនៃលំយោលកើនឡើង នេះអាចនាំឱ្យបាត់បង់ស្ថេរភាពនៃប្រព័ន្ធ និងការផុតពូជនៃប្រភេទសត្វមួយ។
ការធ្វើឱ្យស៊ីជម្រៅបន្ថែមទៀតនៃទ្រឹស្ដីគណិតវិទ្យានៃអន្តរកម្មរវាងប្រភេទសត្វ ដើរតាមខ្សែបន្ទាត់នៃការលម្អិតអំពីរចនាសម្ព័ន្ធនៃចំនួនប្រជាជនខ្លួនឯង និងគិតគូរពីកត្តាបណ្ដោះអាសន្ន និងលំហ។
អក្សរសិល្ប៍។
Kolmogorov A.N. ការសិក្សាគុណភាពនៃគំរូគណិតវិទ្យានៃសក្ដានុពលប្រជាជន។ // បញ្ហានៃ cybernetics ។ ម។, ១៩៧២, លេខ ៥។
MacArtur R. ការវិភាគក្រាហ្វិកនៃប្រព័ន្ធអេកូឡូស៊ី // ផ្នែកនៃរបាយការណ៍ជីវវិទ្យាសាកលវិទ្យាល័យ Perinceton ។ 1971
A.D. Bazykin "ជីវរូបវិទ្យានៃប្រជាជនអន្តរកម្ម" ។ M., Nauka, 1985 ។
V. Volterra: "ទ្រឹស្តីគណិតវិទ្យានៃការតស៊ូដើម្បីអត្ថិភាព" ។ ម.. វិទ្យាសាស្រ្ត, 1976
ហ្គាស G.F. ការតស៊ូដើម្បីអត្ថិភាព។ បាល់ទីម័រ ឆ្នាំ ១៩៣៤។
ម៉ូដែលកុំព្យូទ័រ "PREDATOR-VICTIM"
Kazachkov Igor Alekseevich ១, Guseva Elena Nikolaevna ២
1 សាកលវិទ្យាល័យបច្ចេកទេសរដ្ឋ Magnitogorsk ដាក់ឈ្មោះតាម។ G.I. ណូសូវ៉ា វិទ្យាស្ថានសំណង់ស្ថាបត្យកម្ម និងសិល្បៈ និស្សិតឆ្នាំទី៥
2 សាកលវិទ្យាល័យបច្ចេកទេសរដ្ឋ Magnitogorsk ដាក់ឈ្មោះតាម។ G.I. ណូសូវ៉ា វិទ្យាស្ថានថាមពល និងប្រព័ន្ធស្វ័យប្រវត្តិ បេក្ខជននៃវិទ្យាសាស្ត្រគរុកោសល្យ សាស្ត្រាចារ្យរងនៃនាយកដ្ឋានព័ត៌មានពាណិជ្ជកម្ម និងបច្ចេកវិទ្យាព័ត៌មាន
ចំណារពន្យល់
អត្ថបទនេះត្រូវបានឧទ្ទិសដល់ទិដ្ឋភាពទូទៅនៃគំរូកុំព្យូទ័រ "អ្នកប្រម៉ាញ់-សត្វព្រៃ"។ ការស្រាវជ្រាវដែលបានធ្វើឡើងបានបង្ហាញថា គំរូបរិស្ថានដើរតួនាទីយ៉ាងធំក្នុងការស្រាវជ្រាវបរិស្ថាន។ បញ្ហានេះមានលក្ខណៈចម្រុះ។
ម៉ូដែលកុំព្យូទ័រ "PREDATOR-VICTIM"
Kazatchkov Igor Alekseevich ១, Guseva Elena Nikolaevna ២
1 សាកលវិទ្យាល័យបច្ចេកទេសរដ្ឋ Nosov Magnitogorsk វិស្វកម្មសំណង់ស៊ីវិល ស្ថាបត្យកម្ម និងសិល្បៈ និស្សិតនៃវគ្គសិក្សាទី 5
2 Nosov Magnitogorsk State Technical University, Power Engineering and Automated Systems Institute, PhD in Pedagogical Science, Associate Professor of Business Computer Science and Information Technologies Department
អរូបី
អត្ថបទនេះផ្តល់នូវទិដ្ឋភាពទូទៅនៃគំរូកុំព្យូទ័រ "predator-victim" ។ ការសិក្សាបានបង្ហាញថា ការក្លែងធ្វើបរិស្ថានដើរតួនាទីយ៉ាងធំក្នុងការសិក្សាអំពីបរិស្ថាន។ បញ្ហានេះមានលក្ខណៈចម្រុះ។
គំរូអេកូឡូស៊ីត្រូវបានប្រើដើម្បីសិក្សាបរិស្ថានរបស់យើង។ គំរូគណិតវិទ្យាត្រូវបានប្រើក្នុងករណីដែលមិនមានបរិស្ថានធម្មជាតិ និងគ្មានវត្ថុធម្មជាតិ វាជួយព្យាករណ៍ឥទ្ធិពលនៃកត្តាផ្សេងៗលើវត្ថុដែលកំពុងសិក្សា។ វិធីសាស្រ្តនេះត្រូវចំណាយពេលលើមុខងារនៃការត្រួតពិនិត្យ សាងសង់ និងបកស្រាយលទ្ធផលដែលទទួលបាន។ ដោយផ្អែកលើទម្រង់បែបនេះ គំរូបរិស្ថានទាក់ទងនឹងការវាយតម្លៃនៃការផ្លាស់ប្តូរបរិស្ថានជុំវិញខ្លួនយើង។
បច្ចុប្បន្ននេះទម្រង់បែបនោះត្រូវបានប្រើដើម្បីសិក្សាពីបរិស្ថានជុំវិញខ្លួនយើង ហើយនៅពេលដែលចាំបាច់ត្រូវសិក្សាផ្នែកណាមួយរបស់វា គំរូគណិតវិទ្យាត្រូវបានប្រើប្រាស់។ គំរូនេះធ្វើឱ្យវាអាចទស្សន៍ទាយឥទ្ធិពលនៃកត្តាមួយចំនួនលើវត្ថុនៃការសិក្សា។ នៅពេលមួយ ប្រភេទ "predator-prey" ត្រូវបានស្នើឡើងដោយអ្នកវិទ្យាសាស្ត្រដូចជា៖ T. Malthus (Malthus 1798, Malthus 1905), Verhulst (Verhulst 1838), Pearl (Pearl 1927, 1930) ក៏ដូចជា A. Lotka ( Lotka 1925, 1927 ) និង V. Volterra (Volterra 1926) គំរូទាំងនេះបង្កើតឡើងវិញនូវរបបលំយោលតាមកាលកំណត់ដែលកើតឡើងជាលទ្ធផលនៃអន្តរកម្មអន្តរកម្មនៅក្នុងធម្មជាតិ។
វិធីសាស្រ្តសំខាន់មួយនៃការយល់ដឹងគឺការធ្វើគំរូ។ បន្ថែមពីលើការពិតដែលថាវាអាចទស្សន៍ទាយការផ្លាស់ប្តូរដែលកើតឡើងនៅក្នុង បរិស្ថាន, ក៏ជួយស្វែងរកវិធីល្អបំផុតដើម្បីដោះស្រាយបញ្ហា។ គំរូគណិតវិទ្យាត្រូវបានប្រើប្រាស់ក្នុងបរិស្ថានវិទ្យាអស់រយៈពេលជាយូរ ដើម្បីបង្កើតគំរូ និងនិន្នាការក្នុងការអភិវឌ្ឍន៍ចំនួនប្រជាជន និងជួយគូសបញ្ជាក់ពីខ្លឹមសារនៃការសង្កេត។ ប្លង់អាចធ្វើជាគំរូ អាកប្បកិរិយា, វត្ថុ។
នៅពេលបង្កើតវត្ថុឡើងវិញក្នុងជីវវិទ្យាគណិតវិទ្យា ការព្យាករណ៍នៃប្រព័ន្ធផ្សេងៗត្រូវបានប្រើប្រាស់ លក្ខណៈបុគ្គលពិសេសនៃប្រព័ន្ធជីវសាស្រ្តត្រូវបានផ្តល់ជូនសម្រាប់៖ រចនាសម្ព័ន្ធផ្ទៃក្នុងរបស់បុគ្គល លក្ខខណ្ឌទ្រទ្រង់ជីវិត ភាពស្ថិតស្ថេរនៃប្រព័ន្ធអេកូឡូស៊ី អរគុណដែលសកម្មភាពសំខាន់នៃប្រព័ន្ធត្រូវបានរក្សាទុក។ .
ការមកដល់នៃគំរូកុំព្យូទ័របានឈានទៅមុខយ៉ាងខ្លាំងនូវសមត្ថភាពស្រាវជ្រាវ។ លទ្ធភាពនៃការអនុវត្តពហុភាគីនៃទម្រង់ពិបាកដែលមិនអនុញ្ញាតឱ្យមានការសិក្សាវិភាគបានលេចចេញនូវទិសដៅថ្មី ក៏ដូចជាការធ្វើគំរូ។
ចូរយើងពិចារណាថាតើវត្ថុគំរូគឺជាអ្វី។ "វត្ថុគឺជាជម្រកបិទជិតដែលអន្តរកម្មរវាងចំនួនជីវសាស្រ្តពីរកើតឡើង: មំសាសីនិងសត្វព្រៃ។ ដំណើរការនៃការលូតលាស់ ការផុតពូជ និងការបន្តពូជកើតឡើងដោយផ្ទាល់លើផ្ទៃនៃជម្រក។ សត្វពាហនៈចិញ្ចឹមលើធនធានដែលមាននៅក្នុងបរិស្ថាន ចំណែកសត្វមំសាសីចិញ្ចឹមលើសត្វព្រៃ។ ក្នុងករណីនេះ ធនធានអាហារូបត្ថម្ភអាចកកើតឡើងវិញ ឬមិនអាចកកើតឡើងវិញបាន។
នៅឆ្នាំ 1931 Vito Volterra ទទួលបានច្បាប់ខាងក្រោមនៃទំនាក់ទំនង predator-prey ។
ច្បាប់នៃវដ្ដតាមកាលកំណត់ - ដំណើរការនៃការបំផ្លិចបំផ្លាញសត្វព្រៃដោយសត្វមំសាសី ជារឿយៗនាំឱ្យមានការប្រែប្រួលតាមកាលកំណត់នៃចំនួនប្រជាជននៃប្រភេទទាំងពីរ អាស្រ័យតែលើអត្រាកំណើននៃសត្វស៊ីសាច់ និងសត្វស្មៅ និងលើសមាមាត្រដំបូងនៃចំនួនរបស់វា។
ច្បាប់នៃការអភិរក្សជាមធ្យម - ភាពសម្បូរបែបជាមធ្យមនៃប្រភេទសត្វនីមួយៗគឺថេរ ដោយមិនគិតពីកម្រិតដំបូងឡើយ ផ្តល់ថាអត្រាជាក់លាក់នៃការកើនឡើងចំនួនប្រជាជន ក៏ដូចជាប្រសិទ្ធភាពនៃការចាប់សត្វគឺថេរ។
ច្បាប់នៃការរំលោភលើតម្លៃមធ្យម - នៅពេលដែលប្រភេទសត្វទាំងពីរត្រូវបានកាត់បន្ថយសមាមាត្រទៅនឹងចំនួនរបស់វា ទំហំប្រជាជនជាមធ្យមនៃសត្វព្រៃកើនឡើង ហើយសត្វមំសាសីមានការថយចុះ។
គំរូ predator-prey គឺជាទំនាក់ទំនងពិសេសរវាង predator និង prey របស់វា ជាលទ្ធផលដែលទាំងពីរទទួលបានអត្ថប្រយោជន៍។ បុគ្គលដែលមានសុខភាពល្អបំផុត និងសម្របខ្លួនបំផុតទៅនឹងលក្ខខណ្ឌបរិស្ថានរស់រានមានជីវិត ពោលគឺឧ។ ទាំងអស់នេះកើតឡើងដោយសារតែការជ្រើសរើសធម្មជាតិ។ នៅក្នុងបរិយាកាសដែលមិនមានឱកាសបន្តពូជ។ សត្វមំសាសីនឹងបំផ្លាញចំនួនប្រជាជននៃសត្វព្រៃ មិនយូរមិនឆាប់ ដែលជាលទ្ធផលរបស់វានឹងត្រូវផុតពូជ»។
មានសារពាង្គកាយរស់នៅជាច្រើននៅលើផែនដី ដែលនៅក្រោមលក្ខខណ្ឌអំណោយផល បង្កើនចំនួនសាច់ញាតិរបស់ពួកគេដល់សមាមាត្រដ៏ធំសម្បើម។ សមត្ថភាពនេះត្រូវបានគេហៅថា: សក្តានុពលជីវសាស្ត្រនៃប្រភេទមួយ, i.e. ការកើនឡើងនៃចំនួនប្រភេទសត្វក្នុងរយៈពេលជាក់លាក់ណាមួយ។ ប្រភេទសត្វនីមួយៗមានសក្ដានុពលជីវសាស្រ្តផ្ទាល់ខ្លួន ជាឧទាហរណ៍ ប្រភេទសត្វធំៗអាចកើនឡើងត្រឹមតែ 1.1 ដងក្នុងមួយឆ្នាំ ចំណែកសារពាង្គកាយនៃប្រភេទសត្វតូចៗដូចជា crustaceans ជាដើម។ អាចបង្កើនរូបរាងរបស់ពួកគេរហូតដល់ 1030 ដង ហើយបាក់តេរីនៅក្នុងចំនួនកាន់តែច្រើន។ ក្នុងករណីណាក៏ដោយ ចំនួនប្រជាជននឹងកើនឡើងជាលំដាប់។
កំណើនប្រជាជនអិចស្ប៉ូណង់ស្យែល គឺជាវឌ្ឍនភាពធរណីមាត្រនៃកំណើនប្រជាជន។ សមត្ថភាពនេះអាចត្រូវបានគេសង្កេតឃើញនៅក្នុងមន្ទីរពិសោធន៍ក្នុងបាក់តេរី និងផ្សិត។ នៅក្នុងលក្ខខណ្ឌដែលមិនមែនជាមន្ទីរពិសោធន៍ ការលូតលាស់អិចស្ប៉ូណង់ស្យែលអាចត្រូវបានគេមើលឃើញនៅក្នុងឧទាហរណ៍នៃកណ្តូប ឬនៅក្នុងប្រភេទសត្វល្អិតផ្សេងទៀត។ ការកើនឡើងនៃចំនួនប្រភេទសត្វបែបនេះអាចត្រូវបានគេសង្កេតឃើញនៅកន្លែងទាំងនោះ ដែលវាស្ទើរតែគ្មានសត្រូវ ហើយមានអាហារច្រើនជាងគ្រប់គ្រាន់។ នៅទីបំផុតការកើនឡើងនៃប្រភេទសត្វ បន្ទាប់ពីចំនួនបានកើនឡើងក្នុងរយៈពេលដ៏ខ្លី កំណើនប្រជាជនបានចាប់ផ្តើមធ្លាក់ចុះ។
ចូរយើងពិចារណាគំរូកុំព្យូទ័រនៃការបន្តពូជថនិកសត្វដោយប្រើគំរូ Lotka-Volterra ជាឧទាហរណ៍មួយ។ អនុញ្ញាតឱ្យ នៅតំបន់ជាក់លាក់មួយ សត្វពីរប្រភេទរស់នៅ៖ ក្តាន់ និងចចក។ គំរូគណិតវិទ្យានៃការផ្លាស់ប្តូរចំនួនប្រជាជននៅក្នុងគំរូថាស-Volterra៖
ចំនួនជនរងគ្រោះដំបូងគឺ xn ចំនួនសត្វមំសាសីគឺ yn ។
ប៉ារ៉ាម៉ែត្រគំរូ៖
P1 - ប្រូបាប៊ីលីតេនៃការជួបជាមួយសត្វមំសាសី
P2 - មេគុណនៃការលូតលាស់របស់សត្វមំសាសីដោយចំណាយលើសត្វព្រៃ
ឃ - អត្រាមរណៈរបស់សត្វមច្ឆា
a គឺជាមេគុណនៃការកើនឡើងចំនួនជនរងគ្រោះ។
នៅក្នុងភារកិច្ចបណ្តុះបណ្តាលតម្លៃដូចខាងក្រោមត្រូវបានកំណត់: ចំនួនសត្វក្តាន់គឺ 500 ចំនួនឆ្កែចចកគឺ 10 អត្រាកំណើននៃសត្វក្តាន់គឺ 0.02 អត្រាកំណើននៃសត្វចចកគឺ 0.1 ប្រូបាប៊ីលីតេនៃការជួបសត្វមំសាសីគឺ 0.0026 អត្រាកំណើននៃសត្វមំសាសីក្នុងការចំណាយនៃសត្វព្រៃគឺ 0,000056 ។ ទិន្នន័យត្រូវបានគណនាសម្រាប់ 203 ឆ្នាំ។
យើងស្វែងយល់ពីឥទ្ធិពល មេគុណនៃការកើនឡើងនៃជនរងគ្រោះសម្រាប់ការអភិវឌ្ឍចំនួនប្រជាជនចំនួនពីរ ប៉ារ៉ាម៉ែត្រដែលនៅសល់នឹងមិនត្រូវបានផ្លាស់ប្តូរ។នៅក្នុងគ្រោងការណ៍ទី 1 ការកើនឡើងនៃចំនួនសត្វព្រៃត្រូវបានគេសង្កេតឃើញហើយបន្ទាប់មកជាមួយនឹងការពន្យារពេលខ្លះការកើនឡើងនៃសត្វមំសាសីត្រូវបានអង្កេត។ បន្ទាប់មកសត្វមំសាសីវាយជនរងគ្រោះចំនួនជនរងគ្រោះធ្លាក់ចុះយ៉ាងខ្លាំង ហើយបន្ទាប់ពីវាចំនួនសត្វមំសាសីថយចុះ (រូបភាពទី 1)។

រូបភាពទី 1. ទំហំប្រជាជនដែលមានអត្រាកំណើតទាបក្នុងចំណោមជនរងគ្រោះ
ចូរយើងវិភាគការផ្លាស់ប្តូរគំរូដោយបង្កើនអត្រាកំណើតរបស់ជនរងគ្រោះ a=0.06។ នៅក្នុងដ្យាក្រាម 2 យើងឃើញដំណើរការលំយោលដែលនាំទៅដល់ការកើនឡើងនៃចំនួនប្រជាជនទាំងពីរតាមពេលវេលា (រូបភាពទី 2)។

រូបភាពទី 2. ទំហំប្រជាជននៅអត្រាកំណើតជាមធ្យមរបស់ជនរងគ្រោះ
ចូរយើងពិចារណាពីរបៀបដែលសក្ដានុពលចំនួនប្រជាជននឹងផ្លាស់ប្តូរជាមួយនឹងតម្លៃខ្ពស់នៃអត្រាកំណើតរបស់ជនរងគ្រោះ a=1.13។ នៅក្នុងរូបភព។ 3 មានការកើនឡើងយ៉ាងខ្លាំងនៃចំនួនប្រជាជនទាំងពីរ បន្ទាប់មកដោយការផុតពូជទាំងសត្វព្រៃ និងសត្វមំសាសី។ នេះកើតឡើងដោយសារតែចំនួនសត្វព្រៃបានកើនឡើងដល់កម្រិតដែលធនធានបានចាប់ផ្តើមអស់ ដែលជាហេតុនាំឱ្យសត្វព្រៃផុតពូជ។ ការផុតពូជរបស់សត្វមំសាសីកើតឡើងដោយសារតែចំនួនសត្វព្រៃមានការថយចុះ ហើយសត្វមំសាសីបានអស់ធនធានដើម្បីរស់។
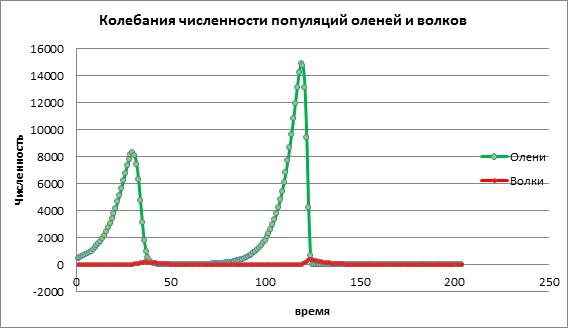
រូបភាពទី 3. ទំហំប្រជាជនដែលមានអត្រាកំណើតខ្ពស់ក្នុងចំណោមជនរងគ្រោះ
ផ្អែកលើការវិភាគទិន្នន័យពិសោធន៍កុំព្យូទ័រ យើងអាចសន្និដ្ឋានបានថា ការធ្វើគំរូកុំព្យូទ័រអនុញ្ញាតឱ្យយើងព្យាករណ៍ពីទំហំប្រជាជន និងសិក្សាពីឥទ្ធិពលនៃកត្តាផ្សេងៗលើសក្ដានុពលចំនួនប្រជាជន។ នៅក្នុងឧទាហរណ៍ខាងលើ យើងបានពិនិត្យមើលគំរូ predator-prey ឥទ្ធិពលនៃអត្រាកំណើតនៃសត្វព្រៃលើចំនួនសត្វក្តាន់ និងចចក។ ការកើនឡើងតិចតួចនៃចំនួនសត្វព្រៃនាំទៅរកការកើនឡើងតិចតួចនៃសត្វព្រៃ ដែលបន្ទាប់ពីរយៈពេលជាក់លាក់ណាមួយត្រូវបានបំផ្លាញដោយសត្វមំសាសី។ការកើនឡើងកម្រិតមធ្យមនៃចំនួនសត្វព្រៃនាំទៅរកការកើនឡើងនៃចំនួនប្រជាជនទាំងពីរ។ ការកើនឡើងខ្ពស់នៃចំនួនសត្វព្រៃ ជាដំបូងនាំឱ្យមានការកើនឡើងយ៉ាងឆាប់រហ័សនៃចំនួនសត្វព្រៃ ដែលវាប៉ះពាល់ដល់ការកើនឡើងនៃចំនួនសត្វមំសាសី ប៉ុន្តែបន្ទាប់មក សត្វមំសាសីដែលមានចំនួនច្រើនបានបំផ្លាញចំនួនសត្វក្តាន់យ៉ាងឆាប់រហ័ស។ ជាលទ្ធផល សត្វទាំងពីរនេះផុតពូជ។
សត្វមំសាសីអាចស៊ីចំណីសត្វ និងសត្វមំសាសីទន់ខ្សោយផងដែរ។ Predators មានអាហារច្រើនប្រភេទ ហើយងាយស្រួលប្តូរពីសត្វព្រៃមួយទៅមួយទៀត ដែលអាចចូលប្រើបានច្រើនជាងមុន។ សត្វឈ្មោលច្រើនតែវាយលុកឈ្មោលខ្សោយ។ តុល្យភាពអេកូឡូស៊ីត្រូវបានរក្សារវាងចំនួនអ្នកប្រម៉ាញ់សត្វព្រៃ។[...]
ប្រសិនបើលំនឹងមិនស្ថិតស្ថេរ (មិនមានវដ្តកំណត់) ឬវដ្ដខាងក្រៅមិនស្ថិតស្ថេរ នោះចំនួននៃប្រភេទសត្វទាំងពីរជួបប្រទះនឹងការប្រែប្រួលខ្លាំង ទុកឱ្យតំបន់ជុំវិញលំនឹង។ លើសពីនេះទៅទៀត, ការចុះខ្សោយយ៉ាងឆាប់រហ័ស (ក្នុងស្ថានភាពដំបូង) កើតឡើងជាមួយនឹងការសម្របខ្លួនទាបនៃ predator, i.e. ជាមួយនឹងអត្រាមរណភាពខ្ពស់របស់វា (បើប្រៀបធៀបទៅនឹងអត្រានៃការបន្តពូជរបស់ជនរងគ្រោះ)។ នេះមានន័យថា សត្វមំសាសីដែលទន់ខ្សោយក្នុងគ្រប់ទិដ្ឋភាពទាំងអស់ មិនរួមចំណែកដល់ស្ថេរភាពនៃប្រព័ន្ធនេះទេ ហើយខ្លួនវាក៏ស្លាប់ទៅវិញ។[...]
សម្ពាធរបស់សត្វមំសាសីគឺខ្លាំងជាពិសេសនៅពេលដែលនៅក្នុងការវិវឌ្ឍន៍របស់ predator-prey តុល្យភាពផ្លាស់ប្តូរឆ្ពោះទៅរក predator ហើយជួរនៃ prey រួមតូច។ ការតស៊ូប្រកួតប្រជែងគឺទាក់ទងយ៉ាងជិតស្និទ្ធទៅនឹងកង្វះខាតធនធានស្បៀង វាក៏អាចជាការតស៊ូផ្ទាល់ផងដែរ ឧទាហរណ៍ សត្វមំសាសីសម្រាប់លំហរជាធនធាន ប៉ុន្តែភាគច្រើនវាគ្រាន់តែជាការផ្លាស់ទីលំនៅរបស់ប្រភេទសត្វដែលមិនមានអាហារគ្រប់គ្រាន់។ ផ្តល់ទឹកដីដោយប្រភេទសត្វដែលមានបរិមាណអាហារគ្រប់គ្រាន់ដូចគ្នា។ នេះគឺជាការប្រកួតប្រជែងគ្នាជាក់លាក់រួចទៅហើយ[...]
 |
ហើយចុងក្រោយនៅក្នុងប្រព័ន្ធ "predator-prey" ដែលបានពិពណ៌នាដោយគំរូ (2.7) ការលេចឡើងនៃអស្ថេរភាពនៃការសាយភាយ (ជាមួយនឹងស្ថេរភាពនៃលំនឹងក្នុងតំបន់) គឺអាចធ្វើទៅបានលុះត្រាតែការស្លាប់ធម្មជាតិរបស់សត្វមំសាសីកើនឡើងជាមួយនឹងកំណើនប្រជាជនរបស់វាលឿនជាង មុខងារលីនេអ៊ែរ និងមុខងារ trophic ខុសគ្នាពី Volterra ឬនៅពេលដែលចំនួនសត្វព្រៃគឺជាចំនួនប្រជាជនប្រភេទ Ollie។[...]
តាមទ្រឹស្តី នៅក្នុងគំរូ "មំសាសីមួយ - សត្វព្រៃពីរ" ការស៊ីស្មៅស្មើៗគ្នា (កង្វះចំណូលចិត្តសម្រាប់ប្រភេទសត្វព្រៃមួយ ឬប្រភេទសត្វផ្សេងទៀត) អាចជះឥទ្ធិពលដល់ការរួមរស់ជាមួយគ្នានៃប្រភេទសត្វព្រៃតែប៉ុណ្ណោះ នៅកន្លែងដែលមានលំនឹងដែលមានសក្តានុពលរួចហើយ។ ភាពចម្រុះអាចកើនឡើងតែនៅក្រោមលក្ខខណ្ឌដែលប្រភេទសត្វដែលមានសមត្ថភាពប្រកួតប្រជែងតិចជាង មានអត្រាកំណើនប្រជាជនខ្ពស់ជាងប្រភេទសត្វដែលលេចធ្លោ។ នេះអនុញ្ញាតឱ្យយើងយល់ពីស្ថានភាពនៅពេលដែលការស៊ីស្មៅឯកសណ្ឋាននាំឱ្យមានការកើនឡើងនៃភាពចម្រុះនៃប្រភេទរុក្ខជាតិ ដែលចំនួនដ៏ច្រើននៃប្រភេទដែលត្រូវបានជ្រើសរើសសម្រាប់ការបន្តពូជយ៉ាងឆាប់រហ័សរួមជាមួយនឹងប្រភេទសត្វដែលការវិវត្តន៍មានគោលបំណងបង្កើនការប្រកួតប្រជែង [... ]
ស្រដៀងគ្នានេះដែរ ការជ្រើសរើសសត្វព្រៃដែលពឹងផ្អែកលើដង់ស៊ីតេអាចនាំទៅរកលំនឹងស្ថិរភាពនៅក្នុងគំរូទ្រឹស្តីនៃប្រភេទសត្វដែលប្រកួតប្រជែងពីរដែលមិនមានលំនឹងពីមុនមក។ ដើម្បីធ្វើដូច្នេះ សត្វមំសាសីត្រូវតែមានសមត្ថភាពឆ្លើយតបមុខងារ និងជាលេខចំពោះការផ្លាស់ប្តូរដង់ស៊ីតេសត្វ។ ទោះយ៉ាងណាក៏ដោយ វាអាចទៅរួចដែលថាការប្តូរ (ការវាយប្រហារញឹកញាប់មិនសមាមាត្រលើសត្វព្រៃដែលមានច្រើនបំផុត) នឹងមានសារៈសំខាន់ជាង។ តាមការពិត វាត្រូវបានបង្កើតឡើងដែលថាការប្តូរមានឥទ្ធិពលស្ថេរភាពនៅក្នុងប្រព័ន្ធ "មួយ predator - n prey" និងតំណាងឱ្យយន្តការតែមួយគត់ដែលមានសមត្ថភាពធ្វើឱ្យអន្តរកម្មមានស្ថេរភាពនៅក្នុងករណីដែល niches សត្វត្រួតស៊ីគ្នាទាំងស្រុង។ សត្វមំសាសីដែលមិនមានជំនាញអាចដើរតួនាទីបែបនេះបាន។ ចំណូលចិត្តដោយសត្វមំសាសីដែលមានឯកទេសកាន់តែច្រើនសម្រាប់គូប្រជែងលេចធ្លោធ្វើសកម្មភាពដូចគ្នានឹងការប្តូរសត្វមំសាសី ហើយអាចរក្សាស្ថេរភាពអន្តរកម្មទ្រឹស្តីនៅក្នុងគំរូដែលពីមុនមិនមានលំនឹងរវាងប្រភេទសត្វព្រៃ ដែលផ្តល់ភាពពិសេសរបស់ពួកគេក្នុងកម្រិតខ្លះបំបែកចេញ។[...]
ដូចគ្នានេះផងដែរ, សត្វមំសាសី "ខ្លាំងគ្រប់បែបយ៉ាង" មិនធ្វើឱ្យសហគមន៍មានស្ថេរភាពទេពោលគឺឧ។ សម្របខ្លួនបានយ៉ាងល្អទៅនឹងសត្វដែលផ្តល់ឲ្យ និងជាមួយនឹងអត្រាមរណភាពដែលទាក់ទងទាប។ ក្នុងករណីនេះ ប្រព័ន្ធមានវដ្ដដែនកំណត់មិនស្ថិតស្ថេរ ហើយទោះបីជាស្ថេរភាពនៃទីតាំងលំនឹងក៏ដោយ ក៏ degenerates នៅក្នុងបរិយាកាសចៃដន្យ (សត្វមំសាសីស៊ីចំណី ហើយជាលទ្ធផលស្លាប់)។ ស្ថានភាពនេះទាក់ទងទៅនឹងការចុះខ្សោយយឺត។
ដូច្នេះជាមួយនឹងការសម្របខ្លួនដ៏ល្អនៃសត្វមំសាសីនៅក្នុងតំបន់ជុំវិញនៃលំនឹងស្ថិរភាពវដ្តមិនស្ថិតស្ថេរនិងស្ថេរភាពអាចកើតឡើង i.e. អាស្រ័យលើលក្ខខណ្ឌដំបូង ប្រព័ន្ធ "អ្នកប្រម៉ាញ់" មានទំនោរទៅរកលំនឹង ឬ លំយោល ផ្លាស់ទីឆ្ងាយពីវា ឬការប្រែប្រួលថេរនៃចំនួននៃប្រភេទទាំងពីរត្រូវបានបង្កើតឡើងនៅក្នុងបរិវេណនៃលំនឹង។
សារពាង្គកាយដែលត្រូវបានចាត់ថ្នាក់ថាជាសត្វមំសាសីចិញ្ចឹមលើសារពាង្គកាយដទៃទៀត បំផ្លាញចំណីរបស់វា។ ដូច្នេះ ក្នុងចំណោមសារពាង្គកាយមានជីវិត គួរតែបែងចែកប្រព័ន្ធចាត់ថ្នាក់មួយបន្ថែមទៀត ពោលគឺ "សត្វមំសាសី" និង "សត្វព្រៃ"។ ទំនាក់ទំនងរវាងសារពាង្គកាយបែបនេះបានរីកចម្រើនពេញមួយការវិវត្តន៍នៃជីវិតនៅលើភពផែនដីរបស់យើង។ សារពាង្គកាយល្មោភកាមដើរតួជានិយតករធម្មជាតិនៃចំនួនសារពាង្គកាយសត្វព្រៃ។ ការកើនឡើងនៃចំនួន "អ្នកប្រម៉ាញ់" នាំឱ្យមានការថយចុះនៃចំនួន "សត្វល្មូន" នេះកាត់បន្ថយការផ្គត់ផ្គង់អាហារ ("សត្វល្មូន") សម្រាប់ "មំសាសី" ដែលជាទូទៅកំណត់ការថយចុះនៃចំនួន។ នៃ "សត្វព្រៃ" ជាដើម។ ដូច្នេះហើយ នៅក្នុង biocenosis ការប្រែប្រួលនៃចំនួនសត្វមំសាសី និងសត្វព្រៃកើតឡើងជានិច្ច ប៉ុន្តែជាទូទៅ លំនឹងជាក់លាក់មួយត្រូវបានបង្កើតឡើងសម្រាប់រយៈពេលជាក់លាក់មួយនៅក្នុងលក្ខខណ្ឌបរិស្ថានដែលមានស្ថិរភាព។[... ]
ទីបំផុតវាមកដល់តុល្យភាពអេកូឡូស៊ីរវាងសត្វមំសាសី និងចំនួនសត្វព្រៃ។[...]
សម្រាប់មុខងារ trophic នៃប្រភេទទីបី ស្ថានភាពលំនឹងនឹងមានស្ថេរភាព ប្រសិនបើកន្លែងដែល N គឺជាចំណុចបញ្ឆេះនៃមុខងារ (សូមមើលរូបទី 2, គ)។ នេះកើតឡើងដោយសារការពិតដែលថា ក្នុងអំឡុងពេលចន្លោះ មុខងារ trophic គឺ concave ហើយដូច្នេះចំណែកទាក់ទងនៃការប្រើប្រាស់សត្វព្រៃដោយ predator កើនឡើង[...]
អនុញ្ញាតឱ្យ Гг = -Г, i.e. មានសហគមន៍ប្រភេទ "អ្នកប្រម៉ាញ់" ។ ក្នុងករណីនេះ ពាក្យទីមួយក្នុងកន្សោម (7.4) គឺស្មើនឹងសូន្យ ហើយដើម្បីបំពេញលក្ខខណ្ឌនៃស្ថេរភាពក្នុងលក្ខខណ្ឌនៃប្រូបាប៊ីលីតេនៃស្ថានភាពលំនឹង N វាត្រូវបានទាមទារថាពាក្យទីពីរក៏មិនវិជ្ជមានដែរ។[... .]
ដូច្នេះ សម្រាប់សហគមន៍សត្វមំសាសីដែលត្រូវបានពិចារណា យើងអាចសន្និដ្ឋានបានថា ទីតាំងលំនឹងវិជ្ជមានជាទូទៅមានស្ថេរភាព asymptotically ពោលគឺ សម្រាប់ទិន្នន័យដំបូងណាមួយ 1H(0)>0 ការវិវត្តកើតឡើងតាមរបៀបដែល N(7) - ■ K បានផ្តល់ថា N > 0 […]
ដូច្នេះហើយ នៅក្នុងបរិយាកាសដូចគ្នាដែលមិនមានជម្រកសម្រាប់ការបន្តពូជ សត្វមំសាសីឆាប់ឬក្រោយមកបំផ្លាញចំនួនប្រជាជននៃសត្វព្រៃ ហើយក្រោយមកក៏ងាប់ដោយខ្លួនវាផ្ទាល់។ រលកនៃជីវិត” (ការផ្លាស់ប្តូរភាពសម្បូរបែបនៃសត្វមំសាសី និងសត្វព្រៃ) ដើរតាមគ្នាជាមួយនឹងការផ្លាស់ប្តូរដំណាក់កាលថេរ ហើយជាមធ្យម ភាពសម្បូរបែបនៃសត្វមំសាសី និងសត្វព្រៃនៅតែមានប្រមាណក្នុងកម្រិតដូចគ្នា។ រយៈពេលនៃរយៈពេលអាស្រ័យលើអត្រាកំណើននៃប្រភេទទាំងពីរនិងលើប៉ារ៉ាម៉ែត្រដំបូង។ សម្រាប់ចំនួនសត្វព្រៃ ឥទ្ធិពលរបស់សត្វមំសាសីគឺមានភាពវិជ្ជមាន ដោយសារការបន្តពូជច្រើនពេករបស់វានឹងនាំទៅដល់ការដួលរលំនៃចំនួនប្រជាជនរបស់វា។ ម៉្យាងវិញទៀត យន្តការទាំងអស់ដែលការពារការសម្លាប់សត្វព្រៃទាំងស្រុង រួមចំណែកដល់ការរក្សាការផ្គត់ផ្គង់អាហាររបស់សត្វមំសាសី។[...]
ការកែប្រែផ្សេងទៀតអាចជាផលវិបាកនៃអាកប្បកិរិយារបស់សត្វមំសាសី។ ចំនួនសត្វព្រៃដែលសត្វមំសាសីអាចស៊ីបាន។ ពេលវេលាដែលបានផ្តល់ឱ្យមានដែនកំណត់របស់វា។ ឥទ្ធិពលនៃការតិត្ថិភាពនៃសត្វមំសាសីនៅពេលជិតដល់កម្រិតនេះត្រូវបានបង្ហាញនៅក្នុងតារាង។ 2-4, B. អន្តរកម្មដែលបានពិពណ៌នាដោយសមីការ 5 និង 6 អាចមានចំណុចលំនឹងថេរ ឬបង្ហាញពីការប្រែប្រួលនៃវដ្ត។ ទោះជាយ៉ាងណាក៏ដោយ វដ្តបែបនេះគឺខុសពីអ្វីដែលឆ្លុះបញ្ចាំងនៅក្នុងសមីការ Lotka-Volterra 1 និង 2។ វដ្តដែលបង្ហាញដោយសមីការ 5 និង 6 អាចមានអំព្លីទីតថេរ និងដង់ស៊ីតេមធ្យម ដរាបណាឧបករណ៍ផ្ទុកគឺថេរ។ បន្ទាប់ពីការរំខានមួយបានកើតឡើង ពួកគេអាចត្រឡប់ទៅទំហំពីមុន និងដង់ស៊ីតេមធ្យមរបស់ពួកគេ។ វដ្តបែបនេះដែលងើបឡើងវិញពីការរំខានត្រូវបានគេហៅថាវដ្តដែនកំណត់មានស្ថេរភាព។ អន្តរកម្មរវាងទន្សាយ និង lynx អាចត្រូវបានចាត់ទុកថាជាវដ្តដែនកំណត់ដែលមានស្ថេរភាព ប៉ុន្តែវាមិនមែនជាវដ្ដ Lotka-Volterra ទេ។[...]
ចូរយើងពិចារណាពីការកើតឡើងនៃអស្ថេរភាពនៃការសាយភាយនៅក្នុងប្រព័ន្ធ predator-prey ប៉ុន្តែជាដំបូងយើងនឹងសរសេរលក្ខខណ្ឌដែលធានាការកើតឡើងនៃអស្ថេរភាពនៃការសាយភាយនៅក្នុងប្រព័ន្ធ (1.1) ជាមួយ n = 2 ។ វាច្បាស់ណាស់ថាលំនឹង (N, N ) គឺក្នុងស្រុក (ឧ។
ចូរយើងបន្តទៅការបកស្រាយនៃករណីដែលទាក់ទងនឹងការរួមរស់ជាមួយគ្នារយៈពេលវែងនៃសត្វមំសាសី និងសត្វព្រៃ។ វាច្បាស់ណាស់ថានៅក្នុងការអវត្ដមាននៃវដ្តកំណត់ លំនឹងស្ថិរភាពនឹងឆ្លើយតបនៅក្នុងបរិយាកាសចៃដន្យចំពោះការប្រែប្រួលចំនួនប្រជាជន ហើយទំហំរបស់ពួកគេនឹងសមាមាត្រទៅនឹងការបែកខ្ញែកនៃការរំខាន។ បាតុភូតនេះនឹងកើតឡើង ប្រសិនបើសត្វមំសាសីមានអត្រាមរណៈដែលទាក់ទងគ្នាខ្ពស់ ហើយក្នុងពេលជាមួយគ្នានោះ កម្រិតខ្ពស់នៃការសម្របខ្លួនទៅនឹងសត្វព្រៃដែលបានផ្តល់ឱ្យ[...]
ឥឡូវនេះសូមឱ្យយើងពិចារណាពីរបៀបដែលថាមវន្តនៃប្រព័ន្ធផ្លាស់ប្តូរជាមួយនឹងការកើនឡើងនៅក្នុងកាយសម្បទារបស់ predator, i.e. ជាមួយនឹង b ថយចុះពី 1 ទៅ 0។ ប្រសិនបើកាយសម្បទាមានកម្រិតទាបគ្រប់គ្រាន់ នោះមិនមានវដ្តកំណត់ទេ ហើយលំនឹងគឺមិនស្ថិតស្ថេរ។ ជាមួយនឹងការកើនឡើងនៃកាយសម្បទានៅក្នុងបរិវេណនៃលំនឹងនេះ វដ្តមានស្ថេរភាព និងបន្ទាប់មកអស្ថិរភាពខាងក្រៅអាចនឹងលេចឡើង។ អាស្រ័យលើលក្ខខណ្ឌដំបូង (សមាមាត្រនៃសត្វមំសាសី និងជីវម៉ាស) ប្រព័ន្ធអាចបាត់បង់ស្ថេរភាព ពោលគឺឧ។ ចាកចេញពីតំបន់ជុំវិញនៃលំនឹង បើមិនដូច្នោះទេលំយោលមានស្ថេរភាពនឹងត្រូវបានបង្កើតឡើងនៅក្នុងវាតាមពេលវេលា។ ការរីកចម្រើនបន្ថែមទៀតនៅក្នុងកាយសម្បទាធ្វើឱ្យលក្ខណៈលំយោលនៃឥរិយាបថនៃប្រព័ន្ធមិនអាចទៅរួចនោះទេ។ ទោះយ៉ាងណាក៏ដោយនៅពេលដែលខ [... ]
ឧទាហរណ៍នៃមតិប្រតិកម្មអវិជ្ជមាន (ស្ថេរភាព) គឺជាទំនាក់ទំនងរវាងមំសាសី និងសត្វព្រៃ ឬដំណើរការនៃប្រព័ន្ធកាបូនសមុទ្រ (ដំណោះស្រាយ CO2 ក្នុងទឹក៖ CO2 + H2O -> H2CO3) ។ ជាធម្មតា បរិមាណកាបូនឌីអុកស៊ីតដែលរលាយក្នុងទឹកសមុទ្រស្ថិតនៅក្នុងលំនឹងដោយផ្នែកជាមួយនឹងការប្រមូលផ្តុំកាបូនឌីអុកស៊ីតនៅក្នុងបរិយាកាស។ ការកើនឡើងនៃកាបូនឌីអុកស៊ីតក្នុងបរិយាកាសបន្ទាប់ពីការផ្ទុះភ្នំភ្លើងនាំឱ្យមានការកើនឡើងនៃការធ្វើរស្មីសំយោគ និងការស្រូបយករបស់វាដោយប្រព័ន្ធកាបូនមហាសមុទ្រ។ នៅពេលដែលកម្រិតកាបូនឌីអុកស៊ីតនៅក្នុងបរិយាកាសមានការថយចុះ ប្រព័ន្ធកាបូណាតនៃមហាសមុទ្របញ្ចេញឧស្ម័នកាបូនិកចូលទៅក្នុងបរិយាកាស។ ដូច្នេះ កំហាប់កាបូនឌីអុកស៊ីតក្នុងបរិយាកាសមានស្ថេរភាពណាស់។[...]
[ ...]
ដូចដែលបានកត់សម្គាល់ដោយ R. Ricklefs (1979) មានកត្តាដែលរួមចំណែកដល់ស្ថេរភាពនៃទំនាក់ទំនងនៅក្នុងប្រព័ន្ធ "predator-prey": អសមត្ថភាពរបស់ predator វត្តមាននៃធនធានអាហារជំនួសសម្រាប់ predator កាត់បន្ថយការពន្យារពេលក្នុង ប្រតិកម្មរបស់សត្វមំសាសី ក៏ដូចជាការរឹតបន្តឹងផ្នែកបរិស្ថានដែលដាក់ដោយបរិយាកាសខាងក្រៅលើប្រជាជនមួយ ឬមួយផ្សេងទៀត។ អន្តរកម្មរវាងចំនួនសត្វមំសាសី និងសត្វព្រៃគឺមានភាពខុសប្លែកគ្នាខ្លាំង និងស្មុគស្មាញ។ ដូច្នេះ ប្រសិនបើសត្វមំសាសីមានប្រសិទ្ធភាពគ្រប់គ្រាន់ ពួកគេអាចគ្រប់គ្រងដង់ស៊ីតេប្រជាជននៃសត្វព្រៃ ដោយរក្សាវាឱ្យទាបជាងសមត្ថភាពផ្ទុករបស់បរិស្ថាន។ តាមរយៈឥទ្ធិពលដែលពួកវាមានលើចំនួនសត្វព្រៃ សត្វមំសាសីមានឥទ្ធិពលលើការវិវត្តនៃលក្ខណៈផ្សេងៗនៃសត្វព្រៃ ដែលនៅទីបំផុតនាំទៅរកតុល្យភាពអេកូឡូស៊ីរវាងចំនួនសត្វមំសាសី និងសត្វព្រៃ។[...]
ប្រសិនបើលក្ខខណ្ឌណាមួយត្រូវបានបំពេញ៖ 0 1/2 ។ ប្រសិនបើ 6 > 1 (kA [... ]
ស្ថេរភាពនៃ biota និងបរិស្ថានអាស្រ័យតែលើអន្តរកម្មនៃរុក្ខជាតិ - autotrophs និងសារពាង្គកាយ heterotrophic herbivorous ។ សត្វមំសាសីគ្រប់ទំហំមិនអាចរំខានដល់តុល្យភាពអេកូឡូស៊ីនៃសហគមន៍បានទេ ពីព្រោះនៅក្រោមលក្ខខណ្ឌធម្មជាតិ ពួកវាមិនអាចបង្កើនចំនួនរបស់ពួកគេជាមួយនឹងចំនួនសត្វព្រៃថេរបានទេ។ សត្វល្មូនមិនត្រឹមតែត្រូវចល័តដោយខ្លួនឯងប៉ុណ្ណោះទេ ថែមទាំងអាចចិញ្ចឹមបានតែលើសត្វដែលផ្លាស់ទីប៉ុណ្ណោះ។[...]
គ្មានត្រីណាផ្សេងទៀតរីករាលដាលដូចជ្រូកនោះទេ។ នៅក្នុងតំបន់នេសាទមួយចំនួននៅក្នុងអាងស្តុកទឹកដែលឈរ ឬហូរ វាមិនមានសម្ពាធពី pikes ដើម្បីរក្សាតុល្យភាពរវាងសត្វព្រៃ និងសត្វមំសាសីទេ មានតែអាងស្តុកទឹកសិប្បនិម្មិតទំនើបប៉ុណ្ណោះ ដែលត្រី pikes គឺជាត្រីដែលមិនចង់បានដោយសារតែការបង្កាត់ពូជត្រីផ្សេងទៀតនៅទីនោះ។ ពួកគេ។ Pike ត្រូវបានតំណាងយ៉ាងល្អនៅក្នុងពិភពលោក។ ពួកគេត្រូវបានចាប់នៅទូទាំងអឌ្ឍគោលខាងជើងពីសហរដ្ឋអាមេរិកនិងកាណាដាទៅ អាមេរិកខាងជើងឆ្លងកាត់អឺរ៉ុបទៅអាស៊ីខាងជើង។
លទ្ធភាពមួយទៀតនៃការរួមរស់ប្រកបដោយនិរន្តរភាពកើតឡើងនៅទីនេះ ក្នុងជួរតូចចង្អៀតនៃការសម្របខ្លួនដែលទាក់ទងខ្ពស់។ នៅពេលដែលការផ្លាស់ប្តូរទៅរបបមិនស្ថិតស្ថេរជាមួយនឹងសត្វមំសាសី "ល្អ" នោះ វដ្តដែនកំណត់ខាងក្រៅដែលមានស្ថេរភាពអាចកើតឡើង ដែលក្នុងនោះការសាយភាយនៃជីវម៉ាសមានតុល្យភាពដោយការហូរចូលទៅក្នុងប្រព័ន្ធ (ផលិតភាពខ្ពស់នៃសត្វព្រៃ)។ បន្ទាប់មកស្ថានភាពចង់ដឹងចង់ឃើញកើតឡើងនៅពេលដែលប្រូបាប៊ីលីតេបំផុតគឺជាតម្លៃលក្ខណៈពីរនៃទំហំនៃការយោលចៃដន្យ។ ខ្លះកើតឡើងនៅជិតលំនឹង ខ្លះទៀតជិតដល់វដ្តកំណត់ ហើយការផ្លាស់ប្តូរញឹកញាប់ ឬតិចរវាងរបបទាំងនេះអាចធ្វើទៅបាន។
ចំនួនប្រជាជនសម្មតិកម្មដែលមានឥរិយាបទយោងទៅតាមវ៉ិចទ័រនៅក្នុងរូបភព។ 10.11 A ត្រូវបានបង្ហាញក្នុងរូប។ 10.11,-B ដោយប្រើក្រាហ្វដែលបង្ហាញពីឌីណាមិកនៃសមាមាត្រនៃចំនួនសត្វមំសាសី និងសត្វព្រៃ និងក្នុងរូបភព។ 10.11.5 ក្នុងទម្រង់ជាក្រាហ្វនៃឌីណាមិកនៃភាពសម្បូរបែបនៃសត្វមំសាសី និងសត្វព្រៃតាមពេលវេលា។ នៅក្នុងចំនួនសត្វព្រៃ នៅពេលដែលវាផ្លាស់ទីពីលំនឹងដង់ស៊ីតេទាបទៅលំនឹងដង់ស៊ីតេខ្ពស់ ហើយត្រលប់មកវិញ "ការផ្ទុះ" នៃចំនួនកើតឡើង។ ហើយការផ្ទុះជាលេខនេះមិនមែនជាលទ្ធផលនៃការផ្លាស់ប្តូរដែលបញ្ចេញសំឡេងស្មើគ្នាក្នុងបរិស្ថាននោះទេ។ ផ្ទុយទៅវិញ ការផ្លាស់ប្តូរលេខនេះត្រូវបានបង្កើតឡើងដោយឥទ្ធិពលខ្លួនវា (ជាមួយនឹងកម្រិតតូចមួយនៃ "សំលេងរំខាន" នៅក្នុងបរិស្ថាន) ហើយជាពិសេស វាឆ្លុះបញ្ចាំងពីអត្ថិភាពនៃស្ថានភាពលំនឹងជាច្រើន។ ហេតុផលស្រដៀងគ្នាអាចត្រូវបានប្រើដើម្បីពន្យល់ករណីស្មុគស្មាញបន្ថែមទៀតនៃសក្ដានុពលចំនួនប្រជាជននៅក្នុងចំនួនប្រជាជនធម្មជាតិ។[...]
ទ្រព្យសម្បត្តិដ៏សំខាន់បំផុតនៃប្រព័ន្ធអេកូគឺស្ថិរភាពរបស់វា តុល្យភាពនៃការផ្លាស់ប្តូរ និងដំណើរការដែលកើតឡើងនៅក្នុងវា។ សមត្ថភាពរបស់ប្រជាជន ឬប្រព័ន្ធអេកូឡូស៊ីក្នុងការរក្សាលំនឹងថាមវន្តដែលមានស្ថេរភាពក្នុងការផ្លាស់ប្តូរលក្ខខណ្ឌបរិស្ថានត្រូវបានគេហៅថា homeostasis (homoios - ដូចគ្នា, ស្រដៀងគ្នា; stasis - រដ្ឋ) ។ Homeostasis គឺផ្អែកលើគោលការណ៍នៃមតិកែលម្អ។ ដើម្បីរក្សាតុល្យភាពនៅក្នុងធម្មជាតិ មិនចាំបាច់មានការគ្រប់គ្រងខាងក្រៅទេ។ ឧទាហរណ៏នៃ homeostasis គឺជាប្រព័ន្ធរង "predator-prey" ដែលក្នុងនោះដង់ស៊ីតេប្រជាជននៃ predator និង prey ត្រូវបានគ្រប់គ្រង។[...]
ប្រព័ន្ធអេកូឡូស៊ីធម្មជាតិ (biogeocenosis) ដំណើរការប្រកបដោយស្ថេរភាពជាមួយនឹងអន្តរកម្មថេរនៃធាតុរបស់វា ចរាចរនៃសារធាតុ ការផ្ទេរសារធាតុគីមី ថាមពល ហ្សែន និងថាមពលផ្សេងទៀត និងព័ត៌មានតាមរយៈខ្សែសង្វាក់។ យោងតាមគោលការណ៍លំនឹង ប្រព័ន្ធធម្មជាតិណាមួយដែលមានលំហូរនៃថាមពល និងព័ត៌មានឆ្លងកាត់វាមានទំនោរនឹងបង្កើតស្ថានភាពស្ថិរភាព។ ទន្ទឹមនឹងនេះ ស្ថិរភាពនៃប្រព័ន្ធអេកូឡូស៊ីត្រូវបានធានាដោយស្វ័យប្រវត្តិតាមរយៈយន្តការផ្តល់មតិ។ មតិកែលម្អរួមមានការប្រើប្រាស់ទិន្នន័យដែលទទួលបានពីសមាសធាតុដែលបានគ្រប់គ្រងនៃប្រព័ន្ធអេកូឡូស៊ី ដើម្បីធ្វើការកែតម្រូវដំណើរការដោយសមាសភាគគ្រប់គ្រង។ ទំនាក់ទំនង "អ្នកប្រម៉ាញ់"-"អ្នកប្រម៉ាញ់" ដែលបានពិភាក្សាខាងលើក្នុងបរិបទនេះ អាចត្រូវបានពិពណ៌នាលម្អិតបន្ថែមទៀត។ បាទ នៅក្នុងប្រព័ន្ធអេកូទឹក ត្រីមំសាសី(pike នៅក្នុងស្រះ) បរិភោគប្រភេទសត្វផ្សេងទៀតនៃត្រី (crucian carp); ប្រសិនបើចំនួននៃត្រីគល់រាំង crucian កើនឡើង នេះគឺជាឧទាហរណ៍នៃមតិស្ថាបនាវិជ្ជមាន។ pike, ចិញ្ចឹមនៅលើ carp crucian, កាត់បន្ថយចំនួនរបស់វា - នេះគឺជាឧទាហរណ៍នៃមតិប្រតិកម្មអវិជ្ជមាន; នៅពេលដែលចំនួនសត្វមំសាសីកើនឡើង ចំនួនជនរងគ្រោះថយចុះ ហើយសត្វមំសាសីដែលជួបប្រទះការខ្វះខាតអាហារ ក៏កាត់បន្ថយកំណើនចំនួនប្រជាជនរបស់វាផងដែរ។ នៅទីបញ្ចប់នៅក្នុងស្រះនៅក្នុងសំណួរលំនឹងថាមវន្តត្រូវបានបង្កើតឡើងនៅក្នុងចំនួននៃត្រីគល់រាំង pike និង crucian ។ លំនឹងត្រូវបានរក្សាជានិច្ច ដែលនឹងមិនរាប់បញ្ចូលការបាត់ខ្លួននៃតំណភ្ជាប់ណាមួយនៅក្នុងខ្សែសង្វាក់ trophic (រូបភាព 64)។[...]
ចូរបន្តឆ្ពោះទៅរកការទូទៅដ៏សំខាន់បំផុត ពោលគឺ អន្តរកម្មអវិជ្ជមានកាន់តែគួរឱ្យកត់សម្គាល់ក្នុងរយៈពេលមួយ ប្រសិនបើប្រព័ន្ធអេកូមានស្ថេរភាពគ្រប់គ្រាន់ ហើយរចនាសម្ព័ន្ធលំហរបស់វាអនុញ្ញាតឱ្យមានការសម្របខ្លួនទៅវិញទៅមកនៃចំនួនប្រជាជន។ នៅក្នុងប្រព័ន្ធគំរូដូចជា predator-preyដែលត្រូវបានពិពណ៌នាដោយសមីការ Lotka-Volterra ប្រសិនបើពាក្យបន្ថែមមិនត្រូវបានណែនាំទៅក្នុងសមីការ កំណត់លក្ខណៈនៃសកម្មភាពនៃកត្តាកំណត់ដោយខ្លួនឯងនៃចំនួន នោះលំយោលកើតឡើងជាបន្តបន្ទាប់ ហើយមិនស្លាប់ទេ (សូមមើល Lewontin, 1969)។ Pimentel (1968; សូមមើលផងដែរ Pimentel and Stone, 1968) បានបង្ហាញដោយពិសោធន៍ថាពាក្យបន្ថែមបែបនេះអាចឆ្លុះបញ្ចាំងពីការសម្របខ្លួនទៅវិញទៅមក ឬហ្សែន មតិកែលម្អ. នៅពេលដែលវប្បធម៌ថ្មីត្រូវបានបង្កើតឡើងពីបុគ្គលដែលធ្លាប់រួមរស់ជាមួយគ្នាអស់រយៈពេលពីរឆ្នាំនៅក្នុងវប្បធម៌ដែលចំនួនរបស់ពួកគេមានការប្រែប្រួលយ៉ាងសំខាន់ វាបានប្រែក្លាយថាពួកគេបានបង្កើតប្រព័ន្ធអេកូឡូស៊ី homeostasis ដែលប្រជាជននីមួយៗត្រូវបាន "គាបសង្កត់" ដោយគ្នាទៅវិញទៅមក។ ដល់កម្រិតមួយដែលវាអាចទៅរួច ដែលការរួមរស់របស់ពួកគេនៅក្នុងលំនឹងដែលមានស្ថេរភាពជាងមុន។
គំរូស្ថានភាព Predator-prey
ចូរយើងពិចារណាអំពីគំរូគណិតវិទ្យានៃសក្ដានុពលនៃការរួមរស់ជាមួយគ្នានៃប្រភេទជីវសាស្រ្តពីរ (ចំនួនប្រជាជន) ដែលធ្វើអន្តរកម្មគ្នាទៅវិញទៅមកតាមប្រភេទ "សត្វឈ្មោល" (ចចក និងទន្សាយ pikes និង carp crucian ។ល។) ដែលហៅថាគំរូ Volter-Lotka . វាត្រូវបានទទួលជាលើកដំបូងដោយ A. Lotka (1925) ហើយបន្តិចក្រោយមក និងដោយឯករាជ្យពី Lotka គំរូស្រដៀងគ្នា និងស្មុគស្មាញជាងនេះ ត្រូវបានបង្កើតឡើងដោយគណិតវិទូជនជាតិអ៊ីតាលី V. Volterra (1926) ដែលការងាររបស់គាត់ពិតជាបានដាក់មូលដ្ឋានគ្រឹះនៃអ្វីដែលគេហៅថា បរិស្ថានវិទ្យាគណិតវិទ្យា។
សូមឱ្យមានប្រភេទជីវសាស្រ្តពីរដែលរស់នៅជាមួយគ្នាក្នុងបរិយាកាសដាច់ស្រយាល។ នេះសន្មត់ថា:
- 1. ជនរងគ្រោះអាចរកអាហារបានគ្រប់គ្រាន់ដើម្បីរស់។
- 2. រាល់ពេលដែលសត្វពាហនៈជួបនឹងសត្វមំសាសី ជននោះសម្លាប់ជនរងគ្រោះ។
សម្រាប់ភាពច្បាស់លាស់ យើងនឹងហៅពួកវាថា crucian carp និង pike ។ អនុញ្ញាតឱ្យ
ស្ថានភាពនៃប្រព័ន្ធត្រូវបានកំណត់ដោយបរិមាណ x(t)និង y(t)- ចំនួននៃ carp crucian និង pike ក្នុងពេលតែមួយ ជីដើម្បីទទួលបានសមីការគណិតវិទ្យាដែលពិពណ៌នាអំពីឌីណាមិក (ការផ្លាស់ប្តូរតាមពេលវេលា) នៃចំនួនប្រជាជន យើងបន្តដូចខាងក្រោម។
ដូចនៅក្នុងគំរូកំណើនប្រជាជនពីមុន (សូមមើលផ្នែក 1.1) សម្រាប់ជនរងគ្រោះ យើងមានសមីការ
កន្លែងណា ក> 0 (អត្រាកំណើតលើសពីអត្រាស្លាប់)
មេគុណ កការកើនឡើងនៃសត្វព្រៃអាស្រ័យទៅលើចំនួនសត្វមំសាសី (ថយចុះជាមួយនឹងការកើនឡើងរបស់វា)។ ក្នុងករណីសាមញ្ញបំផុត។ a- a - fjy (a> 0, p> 0) ។បន្ទាប់មកសម្រាប់ទំហំនៃចំនួនសត្វព្រៃ យើងមានសមីការឌីផេរ៉ង់ស្យែល
សម្រាប់ចំនួនសត្វមំសាសី យើងមានសមីការ

កន្លែងណា ខ>0 (អត្រាមរណៈលើសពីអត្រាកំណើត)។
មេគុណ ខការថយចុះនៃមំសាសីត្រូវបានកាត់បន្ថយប្រសិនបើមានសត្វឈ្មោលដើម្បីចិញ្ចឹម។ ក្នុងករណីសាមញ្ញបំផុតយើងអាចយកបាន។ b - y -Sx (y > 0, ស> 0)។ បន្ទាប់មកសម្រាប់ទំហំនៃចំនួនប្រជាជន predator យើងទទួលបានសមីការឌីផេរ៉ង់ស្យែល

ដូច្នេះ សមីការ (1.5) និង (1.6) តំណាងឱ្យគំរូគណិតវិទ្យានៃបញ្ហានៃអន្តរកម្មប្រជាជនដែលកំពុងពិចារណា។ នៅក្នុងគំរូនេះអថេរ x, yគឺជាស្ថានភាពនៃប្រព័ន្ធ ហើយមេគុណកំណត់លក្ខណៈរចនាសម្ព័ន្ធរបស់វា។ ប្រព័ន្ធ nonlinear (1.5), (1.6) គឺជាគំរូ Voltaire-Lotka ។
សមីការ (1.5) និង (1.6) គួរតែត្រូវបានបំពេញបន្ថែមជាមួយនឹងលក្ខខណ្ឌដំបូង - តម្លៃដែលបានផ្តល់ឱ្យនៃចំនួនប្រជាជនដំបូង។
ឥឡូវនេះ ចូរយើងវិភាគគំរូគណិតវិទ្យាដែលបានសាងសង់។
ចូរយើងបង្កើតទម្រង់ដំណាក់កាលនៃប្រព័ន្ធ (1.5), (1.6) (ក្នុងន័យនៃបញ្ហា X> 0, v >0) ។ ការបែងចែកសមីការ (1.5) ដោយសមីការ (1.6) យើងទទួលបានសមីការជាមួយនឹងអថេរដែលអាចបំបែកបាន
ដោយប្រើសមីការនេះយើងមាន
ទំនាក់ទំនង (1.7) ផ្តល់សមីការនៃគន្លងដំណាក់កាលក្នុងទម្រង់មិនច្បាស់លាស់។ ប្រព័ន្ធ (1.5), (1.6) មានស្ថានភាពស្ថានីដែលបានកំណត់ពី

ពីសមីការ (1.8) យើងទទួលបាន (ចាប់តាំងពី l * ច 0, y* ច 0)
សមភាព (១.៩) កំណត់ទីតាំងលំនឹង (ចំណុច អំពី)(រូបភាព 1.6) ។

ទិសដៅនៃចលនាតាមបណ្តោយគន្លងដំណាក់កាលអាចត្រូវបានកំណត់ពីការពិចារណាបែបនេះ។ សូមឱ្យមាន crucians តិចតួច។ g.e. x ~ 0 បន្ទាប់មកពីសមីការ (1.6) y
គន្លងដំណាក់កាលទាំងអស់ (លើកលែងតែចំណុច 0) ខ្សែកោងបិទជិតគ្របដណ្តប់ទីតាំងលំនឹង។ ស្ថានភាពលំនឹងត្រូវគ្នាទៅនឹងចំនួនថេរនៃ x" និង y" នៃត្រីគល់រាំង crucian និង pike ។ ត្រីគល់រាំង Crucian គុណ, pike ស៊ីពួកវា, ស្លាប់ចេញ, ប៉ុន្តែចំនួននៃពួកគេនិងផ្សេងទៀតមិនផ្លាស់ប្តូរ។ "គន្លងដំណាក់កាលបិទត្រូវគ្នាទៅនឹងការផ្លាស់ប្តូរតាមកាលកំណត់នៃចំនួនត្រីគល់រាំង និង pike ។ ម្យ៉ាងទៀតគន្លងដែលចំនុចដំណាក់កាលផ្លាស់ទីអាស្រ័យលើលក្ខខណ្ឌដំបូង។ ចូរយើងពិចារណាពីរបៀបដែលរដ្ឋផ្លាស់ប្តូរតាមគន្លងដំណាក់កាល។ នៅក្នុងទីតាំង ក(រូបភាព 1.6) ។ មានត្រីគល់រាំង crucian តិចតួចនៅទីនេះ pike ជាច្រើន; pikes មិនមានអ្វីត្រូវបានគេបរិភោគហើយពួកគេបន្តិចម្តងស្លាប់ចេញនិងស្ទើរតែ
បាត់ទាំងស្រុង។ ប៉ុន្តែចំនួននៃត្រីគល់រាំង crucian ក៏ថយចុះស្ទើរតែដល់សូន្យ
តែក្រោយមកនៅពេលដែលមាន pike តិចជាង នៅចំនួននៃត្រីគល់រាំង crucian ចាប់ផ្តើមកើនឡើង; អត្រាកំណើនរបស់ពួកគេកើនឡើង ហើយចំនួនរបស់ពួកគេកើនឡើង - វាកើតឡើងរហូតដល់ចំណុច INប៉ុន្តែការកើនឡើងនៃចំនួនត្រីគល់រាំងដែលនាំឱ្យមានការថយចុះនៃដំណើរការនៃការផុតពូជនៃ shuk ហើយចំនួនរបស់ពួកគេចាប់ផ្តើមកើនឡើង (មានអាហារកាន់តែច្រើន) - គ្រោង ព្រះអាទិត្យ។បន្ទាប់មកមានជ្រូកច្រើន ពួកគេស៊ីត្រីគល់រាំង ហើយស៊ីវាស្ទើរតែទាំងអស់ (ផ្នែក ស៊ីឌី) ។បន្ទាប់ពីនេះ pike ចាប់ផ្តើមងាប់ម្តងទៀតហើយដំណើរការម្តងទៀតជាមួយនឹងរយៈពេលប្រហែល 5-7 ឆ្នាំ។ នៅក្នុងរូបភព។ 1.7 ខ្សែកោងដែលបានសាងសង់ប្រកបដោយគុណភាពនៃការផ្លាស់ប្តូរចំនួននៃ carp crucian និង pike អាស្រ័យលើពេលវេលា។ អតិបរមានៃខ្សែកោងឆ្លាស់គ្នា ហើយចំនួនអតិបរមានៃ pikes យឺតជាងចំនួនអតិបរមានៃ carp crucian ។

ឥរិយាបថនេះគឺជារឿងធម្មតាសម្រាប់ប្រព័ន្ធ predator-prey ផ្សេងៗ។ ឥឡូវនេះ ចូរយើងបកស្រាយលទ្ធផលដែលទទួលបាន។
ទោះបីជាការពិតដែលថាគំរូដែលបានពិចារណាគឺសាមញ្ញបំផុតហើយការពិតអ្វីគ្រប់យ៉ាងកើតឡើងកាន់តែស្មុគស្មាញវាធ្វើឱ្យវាអាចពន្យល់ពីអាថ៌កំបាំងមួយចំនួនដែលមាននៅក្នុងធម្មជាតិ។ រឿងរ៉ាវរបស់អ្នកនេសាទអំពីសម័យកាលដែល "កូនជ្រូកលោតចូលក្នុងដៃ" គឺអាចយល់បាន ភាពញឹកញាប់នៃជំងឺរ៉ាំរ៉ៃ។ល។
សូមកត់សម្គាល់មួយបន្ថែមទៀត ការសន្និដ្ឋានគួរឱ្យចាប់អារម្មណ៍ដែលអាចត្រូវបានធ្វើឡើងពីរូបភព។ ១.៦. ប្រសិនបើនៅចំណុច រមានការចាប់ pikes យ៉ាងឆាប់រហ័ស (នៅក្នុងវាក្យស័ព្ទផ្សេងទៀត - ការបាញ់សត្វចចក) បន្ទាប់មកប្រព័ន្ធ "លោត" ដល់ចំណុច សំណួរហើយចលនាបន្ថែមទៀតកើតឡើងតាមគន្លងបិទជិតនៃទំហំតូចជាង ដែលត្រូវបានរំពឹងទុកដោយវិចារណញាណ។ ប្រសិនបើយើងកាត់បន្ថយចំនួន pike នៅចំណុចមួយ។ Rបន្ទាប់មកប្រព័ន្ធនឹងទៅដល់ចំណុច សហើយចលនាបន្ថែមទៀតនឹងកើតឡើងតាមគន្លង ទំហំធំជាង. ទំហំនៃលំយោលនឹងកើនឡើង។ នេះគឺផ្ទុយពីវិចារណញាណ ប៉ុន្តែវាពន្យល់យ៉ាងច្បាស់អំពីបាតុភូតនេះ៖ ជាលទ្ធផលនៃការបាញ់សត្វចចក ចំនួនរបស់ពួកគេកើនឡើងតាមពេលវេលា។ ដូច្នេះជម្រើសនៃពេលនៃការបាញ់ប្រហារមានសារៈសំខាន់ក្នុងករណីនេះ។
ឧបមាថាចំនួនសត្វល្អិតពីរ (ឧទាហរណ៍ aphid និង ladybug ដែលស៊ី aphids) គឺស្ថិតនៅក្នុងលំនឹងធម្មជាតិ x-x*,y=y*(ចំណុច អំពីនៅក្នុងរូបភព។ ១.៦)។ ពិចារណាពីប្រសិទ្ធភាពនៃការប្រើប្រាស់ថ្នាំសម្លាប់សត្វល្អិតតែមួយ x> 0 នៃជនរងគ្រោះនិង y > 0 នៃសត្វមំសាសីដោយមិនបំផ្លាញវាទាំងស្រុង។ ការថយចុះនៃចំនួនប្រជាជនទាំងពីរនាំឱ្យមានការពិតដែលថាចំណុចតំណាងពីមុខតំណែង អំពីនឹង "លោត" ខិតទៅជិតប្រភពដើមនៃកូអរដោនេ, ដែលជាកន្លែងដែល x > 0, y 0 (រូបទី 1.6) វាកើតឡើងថាជាលទ្ធផលនៃសកម្មភាពនៃថ្នាំសំលាប់សត្វល្អិតដែលត្រូវបានរចនាឡើងដើម្បីបំផ្លាញជនរងគ្រោះ ( aphids ) ចំនួនជនរងគ្រោះ ( aphids ) កើនឡើង ហើយចំនួនសត្វមំសាសី ( សត្វខ្លាឃ្មុំ) ថយចុះ។ វាប្រែថាចំនួននៃមំសាសីអាចក្លាយជាតិចតួចដែលពួកវានឹងប្រឈមមុខនឹងការផុតពូជទាំងស្រុងសម្រាប់ហេតុផលផ្សេងទៀត (គ្រោះរាំងស្ងួតជំងឺ។ ល។ ) ។ ដូច្នេះ ការប្រើប្រាស់ថ្នាំសម្លាប់សត្វល្អិត (លុះត្រាតែពួកវាបំផ្លាញសត្វល្អិតដែលបង្កគ្រោះថ្នាក់ទាំងស្រុង) ទីបំផុតនាំឲ្យមានការកើនឡើងនៃចំនួនសត្វល្អិតទាំងនោះ ដែលចំនួនត្រូវបានគ្រប់គ្រងដោយសត្វល្អិតដទៃទៀត។ ករណីបែបនេះត្រូវបានពិពណ៌នានៅក្នុងសៀវភៅជីវវិទ្យា។
ជាទូទៅ អត្រាកំណើននៃចំនួនជនរងគ្រោះ កអាស្រ័យលើ "L" និង y: ក= a(x, y) (ដោយសារតែវត្តមានរបស់សត្វមំសាសី និងដោយសារតែការរឹតបន្តឹងអាហារ)។
ជាមួយនឹងការផ្លាស់ប្តូរតូចមួយនៅក្នុងគំរូ (1.5), (1.6) ពាក្យតូចៗត្រូវបានបន្ថែមទៅផ្នែកខាងស្តាំនៃសមីការ (ដោយគិតគូរឧទាហរណ៍ការប្រកួតប្រជែងនៃត្រីគល់រាំង crucian សម្រាប់អាហារនិង pike សម្រាប់ត្រីគល់រាំង crucian)
នៅទីនេះ 0 f.i « ១.
ក្នុងករណីនេះការសន្និដ្ឋានអំពីរយៈពេលនៃដំណើរការ (ការវិលត្រឡប់នៃប្រព័ន្ធទៅសភាពដើមរបស់វា) ដែលមានសុពលភាពសម្រាប់គំរូ (1.5), (1.6) បាត់បង់សុពលភាពរបស់វា។ អាស្រ័យលើប្រភេទនៃវិសោធនកម្មតូច / និង gស្ថានភាពដែលបង្ហាញក្នុងរូបគឺអាចធ្វើទៅបាន។ ១.៨.

ក្នុងករណី (1) ស្ថានភាពលំនឹង អំពីនិរន្តរភាព។ សម្រាប់លក្ខខណ្ឌដំបូងផ្សេងទៀតបន្ទាប់ពីគ្រប់គ្រាន់ ពេលវេលាដ៏អស្ចារ្យនេះគឺជាអ្វីដែលត្រូវបានដំឡើង។
ក្នុងករណី (2) ប្រព័ន្ធ "មានភាពច្របូកច្របល់" ។ ស្ថានភាពស្ថានីមិនស្ថិតស្ថេរ។ ប្រព័ន្ធបែបនេះនៅទីបំផុតធ្លាក់ចូលទៅក្នុងជួរនៃតម្លៃបែបនេះ Xនិង y ដែលគំរូឈប់អនុវត្ត។
ក្នុងករណី (3) នៅក្នុងប្រព័ន្ធដែលមានស្ថានភាពស្ថានីមិនស្ថិតស្ថេរ អំពីរបបតាមកាលកំណត់ត្រូវបានបង្កើតឡើងតាមពេលវេលា។ មិនដូចគំរូដើម (1.5), (1.6) ក្នុងគំរូនេះ របបតាមកាលកំណត់ថេរមិនអាស្រ័យលើលក្ខខណ្ឌដំបូងឡើយ។ ដំបូងគម្លាតតូចពីស្ថានភាពស្ថិរភាព អំពីមិននាំឱ្យមានការប្រែប្រួលតិចតួចនៅជុំវិញនោះទេ។ អំពីដូចនៅក្នុងគំរូ Volterra-Lotka ប៉ុន្តែចំពោះលំយោលនៃទំហំដែលបានកំណត់យ៉ាងល្អ (និងឯករាជ្យនៃភាពតូចនៃគម្លាត) ។
នៅក្នុង និង។ Arnold ហៅម៉ូដែល Volterra-Lotka ថារឹងដោយសារតែ ការផ្លាស់ប្តូរតិចតួចរបស់វាអាចនាំឱ្យមានការសន្និដ្ឋានខុសពីអ្វីដែលបានផ្តល់ឱ្យខាងលើ។ ដើម្បីវិនិច្ឆ័យថាតើស្ថានភាពណាមួយដែលបានបង្ហាញក្នុងរូបភព។ 1.8 ដែលត្រូវបានអនុវត្តនៅក្នុងប្រព័ន្ធនេះគឺចាំបាច់បំផុត។ ព័ត៍មានបន្ថែមអំពីប្រព័ន្ធ (អំពីប្រភេទនៃវិសោធនកម្មតូច/ និង g).
នៅទីនេះ ផ្ទុយទៅនឹង (3.2.1) សញ្ញានៃ (-012) និង (+a2i) គឺខុសគ្នា។ ដូចនៅក្នុងករណីនៃការប្រកួតប្រជែង (ប្រព័ន្ធនៃសមីការ (2.2.1)) ប្រភពដើមនៃកូអរដោនេ (1) សម្រាប់ប្រព័ន្ធនេះគឺជាចំណុចពិសេសនៃប្រភេទ "ថ្នាំងមិនស្ថិតស្ថេរ" ។ ស្ថានភាពស្ថិរភាពបីផ្សេងទៀត៖

អត្ថន័យជីវសាស្រ្តទាមទារបរិមាណវិជ្ជមាន X y x 2. សម្រាប់ការបញ្ចេញមតិ (3.3.4) នេះមានន័យថា
ប្រសិនបើមេគុណនៃការប្រកួតប្រជែង intraspecific នៃ predators ក,22 = 0 លក្ខខណ្ឌ (3.3.5) នាំឱ្យលក្ខខណ្ឌ ai2
ប្រភេទដែលអាចកើតមាននៃរូបភាពដំណាក់កាលសម្រាប់ប្រព័ន្ធសមីការ (3.3.1) ត្រូវបានបង្ហាញនៅក្នុងរូបភព។ ៣.២ ក-គ។ Isoclins នៃតង់ហ្សង់ផ្ដេកគឺជាបន្ទាត់ត្រង់
និង isoclines នៃតង់សង់បញ្ឈរគឺត្រង់
ពីរូបភព។ 3.2 បង្ហាញដូចខាងក្រោម។ ប្រព័ន្ធ predator-prey system (3.3.1) អាចមានទីតាំងលំនឹងស្ថិរភាព ដែលចំនួនសត្វព្រៃបានផុតពូជទាំងស្រុង។ (x = 0) ហើយនៅសល់តែសត្វមំសាសីប៉ុណ្ណោះ (ចំណុចទី 2 ក្នុងរូបភាព 3.26)។ ជាក់ស្តែង ស្ថានភាពបែបនេះអាចដឹងបានលុះត្រាតែបន្ថែមលើប្រភេទនៃជនរងគ្រោះនៅក្នុងសំណួរ។ Xមំសាសី X2 មានប្រភពថាមពលបន្ថែម។ ការពិតនេះត្រូវបានឆ្លុះបញ្ចាំងនៅក្នុងគំរូដោយពាក្យវិជ្ជមាននៅជ្រុងខាងស្តាំនៃសមីការសម្រាប់ xs ។ ចំណុចឯកវចនៈ (1) និង (3) (រូបភាព 3.26) មិនស្ថិតស្ថេរ។ លទ្ធភាពទីពីរគឺជាស្ថានភាពស្ថានីដែលមានស្ថេរភាពដែលក្នុងនោះចំនួនប្រជាជននៃសត្វមំសាសីបានស្លាប់ទាំងស្រុងហើយនៅសល់តែសត្វព្រៃប៉ុណ្ណោះ - ចំណុចមានស្ថេរភាព (3) (រូបភាព 3.2a) ។ នៅទីនេះចំណុចឯកវចនៈ (1) ក៏ជាថ្នាំងមិនស្ថិតស្ថេរផងដែរ។
ជាចុងក្រោយ លទ្ធភាពទីបីគឺការរួមរស់ជាមួយគ្នាប្រកបដោយស្ថិរភាពនៃចំនួនប្រជាជននៃសត្វមំសាសី និងសត្វព្រៃ (រូបភាព 3.2 គ) លេខស្ថានីដែលត្រូវបានបង្ហាញដោយរូបមន្ត (3.3.4) ។ ចូរយើងពិចារណាករណីនេះឱ្យបានលំអិត។
ចូរយើងសន្មត់ថាមេគុណនៃការប្រកួតប្រជែង intraspecific គឺស្មើនឹងសូន្យ (អាយ= 0, i = 1, 2) ។ ចូរយើងសន្មត់ថាសត្វមំសាសីចិញ្ចឹមតែលើសត្វព្រៃប៉ុណ្ណោះ។ Xហើយនៅក្នុងការអវត្ដមានរបស់ពួកគេ ពួកគេស្លាប់ក្នុងអត្រា C2 (ក្នុង (3.3.5) C2
ចូរយើងធ្វើការសិក្សាលម្អិតអំពីគំរូនេះ ដោយប្រើសញ្ញាណដែលទទួលយកយ៉ាងទូលំទូលាយបំផុតនៅក្នុងអក្សរសិល្ប៍។ ការកែលម្អឡើងវិញ

អង្ករ។ ៣.២. ទីតាំងនៃ isoclines សំខាន់នៅលើបញ្ឈរដំណាក់កាលនៃប្រព័ន្ធ Volterra predator-prey សម្រាប់សមាមាត្រផ្សេងគ្នានៃប៉ារ៉ាម៉ែត្រ: ក- អំពី -
ជាមួយខ្ញុំ គ២គ២
1, 3 - មិនស្ថិតស្ថេរ 2 - ចំណុចឯកវចនៈមានស្ថេរភាព; វី -
1, 2, 3 - មិនស្ថិតស្ថេរ, 4 - ស្ថេរភាពឯកវចនៈចំណុចសំខាន់
ប្រព័ន្ធ predator-prey ក្នុងសញ្ញាណទាំងនេះមានទម្រង់៖

យើងនឹងសិក្សាពីលក្ខណៈសម្បត្តិនៃដំណោះស្រាយនៃប្រព័ន្ធ (3.3.6) នៅលើយន្តហោះដំណាក់កាល ន1
បើក2
ប្រព័ន្ធមានដំណោះស្រាយស្ថានីពីរ។ ពួកវាអាចកំណត់បានយ៉ាងងាយស្រួលដោយស្មើផ្នែកខាងស្តាំនៃប្រព័ន្ធទៅសូន្យ។ យើងទទួលបាន: 
ដូច្នេះដំណោះស្រាយស្ថានី៖

ចូរយើងពិនិត្យមើលឱ្យកាន់តែច្បាស់អំពីដំណោះស្រាយទីពីរ។ ចូរយើងស្វែងរកអាំងតេក្រាលដំបូងនៃប្រព័ន្ធ (3.3.6) ដែលមិនមាន t.ចូរគុណសមីការទីមួយដោយ -72 ទីពីរដោយ -71 ហើយបន្ថែមលទ្ធផល។ យើងទទួលបាន:
ឥឡូវតោះចែកសមីការទីមួយដោយ ននិងគុណនឹង € 2 ហើយចែកទីពីរដោយ JV 2 ហើយគុណនឹង អ៊ីតោះបន្ថែមលទ្ធផលម្តងទៀត៖
ការប្រៀបធៀប (៣.៣.៧) និង (៣.៣.៨) យើងនឹងមាន៖

ការរួមបញ្ចូលយើងទទួលបាន៖

នេះគឺជាអាំងតេក្រាលដំបូងដែលចង់បាន។ ដូច្នេះ ប្រព័ន្ធ (3.3.6) មានលក្ខណៈអភិរក្សព្រោះវាមានអាំងតេក្រាលដំបូងនៃចលនា បរិមាណដែលជាមុខងារនៃអថេរប្រព័ន្ធ។ ននិង ន2 និងឯករាជ្យនៃពេលវេលា។ ទ្រព្យសម្បត្តិនេះអនុញ្ញាតឱ្យយើងសាងសង់សម្រាប់ប្រព័ន្ធ Volterra ប្រព័ន្ធនៃគំនិតស្រដៀងគ្នាទៅនឹងមេកានិចស្ថិតិ (សូមមើលជំពូកទី 5) ដែលតម្លៃថាមពលនៃប្រព័ន្ធដែលថេរក្នុងពេលវេលាដើរតួនាទីយ៉ាងសំខាន់។
សម្រាប់រាល់ថេរ ពី > 0 (ដែលត្រូវនឹងទិន្នន័យដំបូងជាក់លាក់) អាំងតេក្រាលត្រូវគ្នាទៅនឹងគន្លងជាក់លាក់មួយនៅលើយន្តហោះ ន1 បើក2 បម្រើជាគន្លងនៃប្រព័ន្ធ (៣.៣.៦)។
ចូរយើងពិចារណា វិធីសាស្រ្តក្រាហ្វិកការសាងសង់គន្លងដែលស្នើឡើងដោយ Volterra ខ្លួនឯង។ បានកត់សម្គាល់ឃើញថា ផ្នែកខាងស្តាំរូបមន្ត (3.3.9) អាស្រ័យតែលើ D g 2 ហើយខាងឆ្វេងអាស្រ័យតែលើ ន.ចូរយើងសម្គាល់
ពី (3.3.9) វាដូចខាងក្រោមរវាង Xនិង យមានទំនាក់ទំនងសមាមាត្រ
នៅក្នុងរូបភព។ 3.3 បង្ហាញ quadrants ដំបូងនៃប្រព័ន្ធកូអរដោណេចំនួនបួន XOY, NOY, ន2 OXនិង D G 1 0 ន2 ដើម្បីឱ្យពួកគេទាំងអស់មានដើមកំណើតរួម។
នៅជ្រុងខាងឆ្វេងខាងលើ (ការ៉េ NOY)ក្រាហ្វនៃអនុគមន៍ (3.3.8) ត្រូវបានគូសវាសនៅខាងស្តាំខាងក្រោម (បួនជ្រុង ន2 OX)- ក្រាហ្វមុខងារ យ.មុខងារទីមួយមាននាទី នី =និងទីពីរ - អតិបរមានៅ ន2 = ?-
ទីបំផុតនៅក្នុង quadrant XOYចូរយើងបង្កើតបន្ទាត់ត្រង់ (3.3.12) សម្រាប់ការជួសជុលមួយចំនួន ជាមួយ។
ចូរសម្គាល់ចំណុច ននៅលើអ័ក្ស បើក. ចំណុចនេះត្រូវគ្នាទៅនឹងតម្លៃជាក់លាក់មួយ។ យ(ន 1) ដែលងាយស្រួលរកដោយគូរកាត់កែង

អង្ករ។ ៣.៣.
តាមរយៈ នរហូតដល់វាប្រសព្វជាមួយខ្សែកោង (3.3.10) (សូមមើលរូប 3.3)។ នៅក្នុងវេន តម្លៃ K(D^) ត្រូវគ្នាទៅនឹងចំណុចជាក់លាក់ M នៅលើបន្ទាត់ យ = cXដូច្នេះហើយតម្លៃខ្លះ X(N) = Y(N)/c,ដែលអាចត្រូវបានរកឃើញដោយការគូរកាត់កែង A.M.និង M.D.តម្លៃដែលបានរកឃើញ (ចំណុចនេះត្រូវបានសម្គាល់នៅក្នុងរូបភាពជាមួយអក្សរ ឃ)ចំណុចពីរត្រូវគ្នា។ រនិង ជីនៅលើខ្សែកោង (3.3.11) ។ ដោយប្រើចំណុចទាំងនេះ គូរកាត់កែង យើងនឹងរកឃើញចំណុចពីរក្នុងពេលតែមួយ អ៊ី"និង អ៊ី", ដេកលើខ្សែកោង (3.3.9) កូអរដោនេរបស់ពួកគេ៖

គូរកាត់កែង A.M.យើងបានឆ្លងកាត់ខ្សែកោង (3.3.10) នៅចំណុចមួយទៀត INចំណុចនេះត្រូវគ្នានឹងរឿងដដែល រនិង សំណួរនៅលើខ្សែកោង (3.3.11) និងដូចគ្នា។ ននិង SCH.សំរបសំរួល នចំណុចនេះអាចត្រូវបានរកឃើញដោយទម្លាក់កាត់កែងពី INក្នុងមួយអ័ក្ស បើកដូច្នេះយើងទទួលបានពិន្ទុ F"និង F" ក៏ដេកនៅលើខ្សែកោង (3.3.9) ។
មកពីចំណុចផ្សេង Nតាមរបៀបដូចគ្នាយើងទទួលបានបួនចំណុចថ្មីដែលស្ថិតនៅលើខ្សែកោង (3.3.9)។ ករណីលើកលែងនឹងក្លាយជាចំណុច នី= ?2/72- ផ្អែកលើវា យើងទទួលបានតែពីរពិន្ទុប៉ុណ្ណោះ៖ TOនិង អិលទាំងនេះនឹងជាចំណុចខាងក្រោម និងខាងលើនៃខ្សែកោង (3.3.9)។
មិនអាចចាប់ផ្តើមពីតម្លៃបានទេ។ ននិងពីតម្លៃ ន2 . ធ្វើដំណើរពី ន2 ទៅខ្សែកោង (3.3.11) បន្ទាប់មកកើនឡើងដល់បន្ទាត់ត្រង់ Y = cX ហើយពីទីនោះឆ្លងកាត់ខ្សែកោង (3.3.10) យើងក៏រកឃើញចំណុចបួននៃខ្សែកោង (3.3.9) ។ ករណីលើកលែងនឹងក្លាយជាចំណុច ទេ =?1/71- ផ្អែកលើវា យើងទទួលបានតែពីរពិន្ទុប៉ុណ្ណោះ៖ ជីនិង TOទាំងនេះនឹងជាចំណុចឆ្វេងបំផុត និងស្ដាំបំផុតនៃខ្សែកោង (3.3.9)។ ដោយការសួរខុសគ្នា ននិង ន2 ហើយដោយទទួលបានចំណុចច្រើនណាស់ដោយភ្ជាប់ពួកវា យើងនឹងសង់ខ្សែកោងប្រហែល (៣.៣.៩)។
ពីការសាងសង់វាច្បាស់ណាស់ថានេះគឺជាខ្សែកោងបិទដែលមាននៅក្នុងខ្លួនវាផ្ទាល់ចំណុច 12 = (?2/721?1/71)” ចាប់ផ្តើមពីទិន្នន័យដំបូងជាក់លាក់។ ន Yu និង N20 ។ យកតម្លៃមួយទៀតនៃ C, i.e. ទិន្នន័យដំបូងផ្សេងទៀត យើងទទួលបានខ្សែកោងបិទមួយផ្សេងទៀតដែលមិនប្រសព្វគ្នាដំបូង ហើយក៏មានចំណុច (?2/721 ?1/71)1 នៅខាងក្នុងខ្លួនវាផងដែរ។ ដូច្នេះ ក្រុមគ្រួសារនៃគន្លង (3.3.9) គឺជាគ្រួសារនៃបន្ទាត់បិទជុំវិញចំណុច 12 (សូមមើលរូប 3.3)។ ចូរយើងសិក្សាពីប្រភេទនៃស្ថេរភាពនៃចំណុចឯកវចនៈនេះដោយប្រើវិធីសាស្ត្រ Lyapunov ។
ចាប់តាំងពីប៉ារ៉ាម៉ែត្រទាំងអស់។ អ៊ី 1, ?2, 71,72 គឺវិជ្ជមាន, រយៈពេល (N[ មានទីតាំងនៅក្នុង quadrant វិជ្ជមាននៃយន្តហោះដំណាក់កាល។ ការកំណត់ប្រព័ន្ធនៅជិតចំណុចនេះផ្តល់ឱ្យ៖

នៅទីនេះ n(t)និង 7i2(N1, ន2 :
សមីការលក្ខណៈនៃប្រព័ន្ធ (៣.៣.១៣)៖

ឫសគល់នៃសមីការនេះគឺជាការស្រមើលស្រមៃសុទ្ធសាធ៖

ដូច្នេះ ការសិក្សាអំពីប្រព័ន្ធបង្ហាញថាគន្លងនៅជិតចំណុចឯកវចនៈត្រូវបានតំណាងដោយពងក្រពើប្រមូលផ្តុំ ហើយចំនុចឯកវចនៈខ្លួនវាគឺជាចំណុចកណ្តាល (រូបភាព 3.4) ។ គំរូ Volterra ដែលកំពុងត្រូវបានពិចារណាក៏មានគន្លងបិទជិតឆ្ងាយពីចំណុចឯកវចនៈ បើទោះបីជារូបរាងគន្លងទាំងនេះខុសពីរាងអេលីបស្យុងក៏ដោយ។ ឥរិយាបទនៃអថេរ នី, ន2 លើសម៉ោងត្រូវបានបង្ហាញនៅក្នុងរូបភព។ ៣.៥.

អង្ករ។ ៣.៤.

អង្ករ។ ៣.៥. ការពឹងផ្អែកលើចំនួនសត្វព្រៃ នខ្ញុំ និង អ្នកប្រមាញ់ ន2 ពីពេលវេលា
ចំណុចឯកវចនៈប្រភេទកណ្តាលមានស្ថេរភាព ប៉ុន្តែមិនមាន asymptotically ។ ចូរយើងបង្ហាញជាមួយឧទាហរណ៍នេះថាតើនេះជាអ្វី។ អនុញ្ញាតឱ្យមានការស្ទាក់ស្ទើរ Ni(t)និង LGgM កើតឡើងតាមរបៀបដែលចំណុចតំណាងផ្លាស់ទីតាមយន្តហោះដំណាក់កាលតាមគន្លងទី 1 (សូមមើលរូប 3.4)។ នៅពេលចំនុចស្ថិតនៅទីតាំង M ចំនួនជាក់លាក់នៃបុគ្គលត្រូវបានបញ្ចូលទៅក្នុងប្រព័ន្ធពីខាងក្រៅ ន 2, ដូចថាចំណុចតំណាងលោតពីចំណុច មចង្អុល A/" បន្ទាប់ពីនេះ ប្រសិនបើប្រព័ន្ធត្រូវបានទុកចោលម្តងទៀត នោះលំយោល។ នីនិង ន2 នឹងកើតឡើងរួចទៅហើយជាមួយនឹងទំហំធំជាងពីមុន ហើយចំណុចតំណាងផ្លាស់ទីតាមគន្លង 2. នេះមានន័យថាលំយោលនៅក្នុងប្រព័ន្ធមិនស្ថិតស្ថេរ៖ ពួកវាផ្លាស់ប្តូរលក្ខណៈរបស់ពួកគេជារៀងរហូតនៅពេលដែល ឥទ្ធិពលខាងក្រៅ. នៅពេលអនាគត យើងនឹងពិចារណាគំរូដែលពិពណ៌នាអំពីរបបលំយោលដែលមានស្ថេរភាព ហើយបង្ហាញថានៅលើយន្តហោះដំណាក់កាលដូចជាចលនាតាមកាលកំណត់ដែលមានស្ថេរភាព asymptotic ត្រូវបានបង្ហាញដោយប្រើវដ្តកំណត់។
នៅក្នុងរូបភព។ រូបភាព 3.6 បង្ហាញពីខ្សែកោងពិសោធន៍ - ការប្រែប្រួលនៃចំនួនសត្វខ្លាឃ្មុំនៅប្រទេសកាណាដា (យោងទៅតាមក្រុមហ៊ុន Hudson's Bay) ។ ខ្សែកោងទាំងនេះត្រូវបានសាងសង់ដោយផ្អែកលើទិន្នន័យអំពីចំនួនស្បែកដែលប្រមូលផល។ រយៈពេលនៃការប្រែប្រួលនៃចំនួនទន្សាយ (សត្វព្រៃ) និង lynxes (អ្នកបរបាញ់) គឺប្រហែលដូចគ្នា ហើយមានរយៈពេលប្រហែល 9-10 ឆ្នាំ។ ក្នុងករណីនេះចំនួនអតិបរមានៃ hares គឺ, ជាក្បួន, នាំមុខចំនួនអតិបរមានៃ lynxes ដោយមួយឆ្នាំ។
រូបរាងនៃខ្សែកោងពិសោធន៍ទាំងនេះគឺមានភាពទៀងទាត់តិចជាងទ្រឹស្ដី។ ទោះជាយ៉ាងណាក៏ដោយក្នុងករណីនេះវាគ្រប់គ្រាន់ហើយដែលគំរូធានានូវភាពចៃដន្យនៃលក្ខណៈសំខាន់ៗបំផុតនៃខ្សែកោងទ្រឹស្តីនិងពិសោធន៍ពោលគឺឧ។ តម្លៃអំព្លីទីត និងការផ្លាស់ប្តូរដំណាក់កាលរវាងការប្រែប្រួលនៃចំនួនសត្វមំសាសី និងសត្វព្រៃ។ គុណវិបត្តិដ៏ធ្ងន់ធ្ងរជាងនេះនៃគំរូ Volterra គឺអស្ថិរភាពនៃដំណោះស្រាយចំពោះប្រព័ន្ធសមីការ។ ជាការពិតណាស់ ដូចដែលបានរៀបរាប់ខាងលើ ការផ្លាស់ប្តូរចៃដន្យណាមួយនៅក្នុងភាពសម្បូរបែបនៃប្រភេទសត្វមួយ ឬប្រភេទផ្សេងទៀតគួរតែនាំទៅរកការផ្លាស់ប្តូរនៃទំហំនៃការយោលនៃប្រភេទទាំងពីរ។ តាមធម្មជាតិនៅក្នុង លក្ខខណ្ឌធម្មជាតិសត្វត្រូវបានទទួលរងនូវឥទ្ធិពលចៃដន្យបែបនេះរាប់មិនអស់។ ដូចដែលអាចមើលឃើញពីខ្សែកោងពិសោធន៍ ទំហំនៃភាពប្រែប្រួលនៃចំនួនប្រភេទសត្វប្រែប្រួលតិចតួចពីមួយឆ្នាំទៅមួយឆ្នាំ។
គំរូ Volterra គឺជាឯកសារយោង (មូលដ្ឋាន) សម្រាប់បរិស្ថានវិទ្យាគណិតវិទ្យាក្នុងកម្រិតដូចគ្នាដែលគំរូលំយោលអាម៉ូនិកគឺជាមូលដ្ឋានសម្រាប់បុរាណ និង មេកានិចកង់ទិច. ការប្រើប្រាស់គំរូនេះ ដោយផ្អែកលើគំនិតសាមញ្ញបំផុតអំពីធម្មជាតិនៃគំរូដែលពិពណ៌នាអំពីឥរិយាបថនៃប្រព័ន្ធ គណិតវិទ្យាសុទ្ធសាធ
ជំពូកទី 3

អង្ករ។ ៣.៦. ខ្សែកោង Kinetic នៃចំនួនសត្វខ្លាឃ្មុំ យោងទៅតាមទិន្នន័យរបស់ក្រុមហ៊ុន Hudson Bay Fur (Seton-Thomson, 1987) តាមរយៈការសន្និដ្ឋានមួយត្រូវបានគូរអំពីលក្ខណៈគុណភាពនៃឥរិយាបទនៃប្រព័ន្ធបែបនេះ - អំពីវត្តមានរបស់ ការប្រែប្រួលចំនួនប្រជាជននៅក្នុងប្រព័ន្ធបែបនេះ។ បើគ្មានការបង្កើតគំរូគណិតវិទ្យា និងប្រើវា ការសន្និដ្ឋានបែបនេះនឹងមិនអាចទៅរួចទេ។
នៅក្នុងអ្វីដែលយើងបានពិភាក្សាខាងលើ ក្នុងទម្រង់សាមញ្ញប្រព័ន្ធ Volterra មានគុណវិបត្តិជាមូលដ្ឋាន និងទំនាក់ទំនងគ្នាពីរ។ អក្សរសិល្ប៍អេកូឡូស៊ី និងគណិតវិទ្យាយ៉ាងទូលំទូលាយត្រូវបានឧទ្ទិសដល់ "ការលុបបំបាត់" របស់ពួកគេ។ ទីមួយ ការដាក់បញ្ចូលកត្តាបន្ថែមណាមួយ ទោះជាតូចក៏ដោយ នៅក្នុងគំរូបានផ្លាស់ប្តូរឥរិយាបថនៃប្រព័ន្ធប្រកបដោយគុណភាព។ គុណវិបត្តិ "ជីវសាស្រ្ត" ទីពីរនៃគំរូគឺថាវាមិនរាប់បញ្ចូលលក្ខណៈសម្បត្តិជាមូលដ្ឋានដែលមាននៅក្នុងចំនួនប្រជាជនណាមួយដែលធ្វើអន្តរកម្មគ្នាដោយយោងទៅតាមគោលការណ៍ predator-prey: ឥទ្ធិពលនៃការតិត្ថិភាពនៃសត្វមំសាសី ធនធានមានកំណត់របស់ predator និង prey សូម្បីតែ ជាមួយនឹងចំនួនសត្វព្រៃច្រើន លទ្ធភាពនៃចំនួនសត្វព្រៃដែលអាចរកបានសម្រាប់សត្វរំពា។ល។
ដើម្បីលុបបំបាត់ការខ្វះខាតទាំងនេះ ការកែប្រែផ្សេងៗនៃប្រព័ន្ធ Volterra ត្រូវបានស្នើឡើងដោយអ្នកនិពន្ធផ្សេងៗគ្នា។ អ្វីដែលគួរឱ្យចាប់អារម្មណ៍បំផុតនៃពួកគេនឹងត្រូវបានពិភាក្សានៅក្នុងផ្នែក 3.5 ។ នៅទីនេះយើងនឹងផ្តោតតែលើគំរូដែលគិតគូរពីការកំណត់ដោយខ្លួនឯងក្នុងកំណើននៃចំនួនប្រជាជនទាំងពីរ។ ឧទាហរណ៍នៃគំរូនេះបង្ហាញយ៉ាងច្បាស់ពីរបៀបដែលធម្មជាតិនៃការសម្រេចចិត្តអាចផ្លាស់ប្តូរនៅពេលដែលប៉ារ៉ាម៉ែត្រប្រព័ន្ធផ្លាស់ប្តូរ។
ដូច្នេះយើងពិចារណាប្រព័ន្ធ

ប្រព័ន្ធ (3.3.15) ខុសពីប្រព័ន្ធដែលបានពិចារណាពីមុន (3.3.6) ដោយវត្តមាននៃលក្ខខណ្ឌនៃទម្រង់ -7 នៅខាងស្តាំដៃនៃសមីការ uNf,
ពាក្យទាំងនេះឆ្លុះបញ្ចាំងពីការពិតដែលថាចំនួនសត្វព្រៃមិនអាចកើនឡើងដោយគ្មានកំណត់សូម្បីតែនៅក្នុងអវត្តមាននៃមំសាសីដោយសារតែធនធានអាហារមានកម្រិត និងទីជម្រកមានកំណត់។ "ការរឹតបន្តឹងដោយខ្លួនឯង" ដូចគ្នាត្រូវបានដាក់លើចំនួនប្រជាជននៃសត្វមំសាសី។
ដើម្បីស្វែងរកចំនួនថេរនៃប្រភេទ iVi និង ន2 ចូរយើងធ្វើសមភាពផ្នែកខាងស្តាំនៃសមីការនៃប្រព័ន្ធ (3.3.15) ទៅសូន្យ។ ដំណោះស្រាយជាមួយនឹងលេខសូន្យនៃមំសាសី ឬសត្វព្រៃនឹងមិនចាប់អារម្មណ៍យើងឥឡូវនេះទេ។ ដូច្នេះសូមពិចារណាប្រព័ន្ធពិជគណិត
សមីការ  ការសម្រេចចិត្តរបស់នាង
ការសម្រេចចិត្តរបស់នាង
ផ្តល់ឱ្យយើងនូវកូអរដោនេនៃចំណុចឯកវចនៈ។ ប៉ារ៉ាម៉ែត្រប្រព័ន្ធនៅទីនេះគួរតែស្ថិតនៅក្រោមលក្ខខណ្ឌដែលលេខស្ថានីគឺវិជ្ជមាន៖ ន> 0 និង ន2 > 0. ឫសគល់នៃសមីការលក្ខណៈនៃប្រព័ន្ធលីនេអ៊ែរក្នុងសង្កាត់នៃចំណុចឯកវចនៈ (៣.៣.១៦)៖

ពីកន្សោមសម្រាប់លេខលក្ខណៈវាច្បាស់ណាស់ថាប្រសិនបើលក្ខខណ្ឌត្រូវបានបំពេញ
បន្ទាប់មកចំនួននៃសត្វមំសាសី និងសត្វព្រៃបានឆ្លងកាត់ការយោលសើមៗតាមពេលវេលា ប្រព័ន្ធមានចំណុចឯកវចនៈមិនសូន្យ និងការផ្តោតអារម្មណ៍ថេរ។ បញ្ឈរដំណាក់កាលនៃប្រព័ន្ធបែបនេះត្រូវបានបង្ហាញនៅក្នុងរូបភព។ ៣.៧ ក.
ចូរយើងសន្មត់ថាប៉ារ៉ាម៉ែត្រនៅក្នុងវិសមភាព (3.3.17) ផ្លាស់ប្តូរតម្លៃរបស់ពួកគេតាមរបៀបដែលលក្ខខណ្ឌ (3.3.17) ក្លាយជាសមភាព។ បន្ទាប់មកលេខលក្ខណៈនៃប្រព័ន្ធ (3.3.15) គឺស្មើគ្នា ហើយចំនុចឯកវចនៈរបស់វានឹងស្ថិតនៅលើព្រំប្រទល់រវាងតំបន់នៃ foci និងថ្នាំងដែលមានស្ថេរភាព។ នៅពេលដែលសញ្ញានៃវិសមភាព (3.3.17) ត្រូវបានបញ្ច្រាសចំណុចឯកវចនៈក្លាយជាថ្នាំងដែលមានស្ថេរភាព។ បញ្ឈរដំណាក់កាលនៃប្រព័ន្ធសម្រាប់ករណីនេះត្រូវបានបង្ហាញនៅក្នុងរូបភព។ ៣.៧៦.
ដូចនៅក្នុងករណីនៃចំនួនប្រជាជនតែមួយ គំរូ stochastic អាចត្រូវបានបង្កើតឡើងសម្រាប់គំរូ (3.3.6) ប៉ុន្តែដំណោះស្រាយច្បាស់លាស់មិនអាចទទួលបានសម្រាប់វាទេ។ ដូច្នេះ យើងនឹងដាក់កម្រិតខ្លួនយើងចំពោះការពិចារណាទូទៅ។ ជាឧទាហរណ៍ ចូរយើងសន្មត់ថា ចំណុចលំនឹងស្ថិតនៅចម្ងាយជាក់លាក់មួយពីអ័ក្សនីមួយៗ។ បន្ទាប់មកសម្រាប់គន្លងដំណាក់កាលដែលតម្លៃនៃ JVj, ន2 នៅតែមានទំហំធំល្មម គំរូកំណត់នឹងពេញចិត្តណាស់។ ប៉ុន្តែប្រសិនបើនៅចំណុចណាមួយ។


អង្ករ។ ៣.៧. ដំណាក់កាលបញ្ឈរនៃប្រព័ន្ធ (3.3.15): ក -នៅពេលដែលទំនាក់ទំនង (3.3.17) រវាងប៉ារ៉ាម៉ែត្រត្រូវបានពេញចិត្ត; ខ- នៅពេលអនុវត្តទំនាក់ទំនងបញ្ច្រាសរវាងប៉ារ៉ាម៉ែត្រ
គន្លងដំណាក់កាល អថេរណាមួយមិនធំខ្លាំងទេ បន្ទាប់មកការប្រែប្រួលចៃដន្យអាចក្លាយជាសំខាន់។ ពួកវានាំទៅរកការពិតដែលថាចំណុចតំណាងផ្លាស់ទីទៅអ័ក្សមួយក្នុងចំណោមអ័ក្សដែលមានន័យថាការផុតពូជនៃប្រភេទសត្វដែលត្រូវគ្នា។ ដូច្នេះគំរូ stochastic ប្រែទៅជាមិនស្ថិតស្ថេរចាប់តាំងពី stochastic "រសាត់" ឆាប់ឬក្រោយមកនាំឱ្យមានការផុតពូជនៃប្រភេទសត្វមួយ។ នៅក្នុងគំរូប្រភេទនេះ សត្វមំសាសីនៅទីបំផុតបានផុតពូជ ទោះដោយចៃដន្យ ឬដោយសារចំនួនសត្វព្រៃរបស់វាត្រូវបានកំចាត់មុនគេ។ គំរូ stochastic នៃប្រព័ន្ធ predator-prey ពន្យល់យ៉ាងល្អអំពីការពិសោធន៍របស់ Gause (Gause, 1934; 2000) ដែលក្នុងនោះ ciliates Paramettum candatumបានបម្រើជាជនរងគ្រោះសម្រាប់ ciliate មួយផ្សេងទៀត ឌីឌីនីញ៉ូម ណាសាតម- សត្វមំសាសី។ ចំនួនលំនឹងដែលរំពឹងទុកយោងទៅតាមសមីការកំណត់ (៣.៣.៦) នៅក្នុងការពិសោធន៍ទាំងនេះមានប្រមាណតែ ៥ នាក់នៃប្រភេទនីមួយៗ ដូច្នេះវាមិនគួរឱ្យភ្ញាក់ផ្អើលទេដែលថានៅក្នុងការពិសោធន៍ម្តងហើយម្តងទៀតនីមួយៗ ទាំងសត្វមំសាសី ឬសត្វព្រៃ (ហើយបន្ទាប់មកសត្វមំសាសី) បានស្លាប់យ៉ាងលឿន។ )
ដូច្នេះការវិភាគនៃគំរូ Volterra នៃអន្តរកម្មនៃប្រភេទសត្វបង្ហាញថា ទោះបីជាមានភាពខុសគ្នានៃប្រភេទនៃឥរិយាបទនៃប្រព័ន្ធបែបនេះក៏ដោយ ក៏មិនអាចមានភាពប្រែប្រួលនៃចំនួននៅក្នុងគំរូនៃប្រភេទសត្វដែលប្រកួតប្រជែងគ្នាបានឡើយ។ នៅក្នុងគំរូ predator-prey លំយោលគ្មានសំណើមលេចឡើងជាលទ្ធផលនៃជម្រើស ទម្រង់ពិសេសសមីការគំរូ (៣.៣.៦)។ ក្នុងករណីនេះ គំរូក្លាយជាមិនរដុប ដែលបង្ហាញពីអវត្តមាននៅក្នុងប្រព័ន្ធនៃយន្តការបែបនេះ ដែលស្វែងរកការរក្សាសភាពរបស់វា។ ទោះជាយ៉ាងណាក៏ដោយការយោលបែបនេះត្រូវបានគេសង្កេតឃើញនៅក្នុងធម្មជាតិនិងការពិសោធន៍។ តម្រូវការសម្រាប់ការពន្យល់តាមទ្រឹស្ដីរបស់ពួកគេបានបម្រើជាហេតុផលមួយសម្រាប់បង្កើតការពិពណ៌នាអំពីគំរូបន្ថែមទៀត ទិដ្ឋភាពទូទៅ. ផ្នែកទី 3.5 ត្រូវបានឧទ្ទិសដល់ការពិចារណាលើគំរូទូទៅបែបនេះ។