ការជ្រើសរើសធម្មជាតិ គឺជាកត្តានាំផ្លូវនៃការវិវត្តន៍។ អរូបី៖ ការវិវត្តន៍ និងការជ្រើសរើសធម្មជាតិ មូលដ្ឋានសម្រាប់ភាពជោគជ័យនៃការវិវត្តន៍គឺ
សៀវភៅសិក្សាសម្រាប់ថ្នាក់ទី ១០-១១
§ 46. ការជ្រើសរើសធម្មជាតិគឺជាកត្តាដឹកនាំនៃការវិវត្តន៍
គុណសម្បត្តិដ៏អស្ចារ្យរបស់ Charles Darwin គឺការរកឃើញនូវតួនាទីនៃការជ្រើសរើសជាកត្តាសំខាន់បំផុតក្នុងដំណើរការវិវត្តន៍។ ដាវីនបានជឿថា ដោយសារការជ្រើសរើសធម្មជាតិ ដំណើរការនៃការអភិរក្ស និងការបន្តពូជដោយអនុគ្រោះនៃសារពាង្គកាយដែលមានលក្ខណៈដែលមានប្រយោជន៍បំផុតក្នុងលក្ខខណ្ឌបរិស្ថានត្រូវបានអនុវត្ត។ ការជ្រើសរើសធម្មជាតិ ដូចដែលដាវីនបានបង្ហាញ គឺជាផលវិបាកនៃការតស៊ូដើម្បីអត្ថិភាព។
ការតស៊ូដើម្បីអត្ថិភាព។ដាវីនបានប្រើគំនិតនេះដើម្បីកំណត់លក្ខណៈនៃទំនាក់ទំនងទាំងមូលរវាងបុគ្គល និងកត្តាបរិស្ថានផ្សេងៗ។ ទំនាក់ទំនងទាំងនេះកំណត់ភាពជោគជ័យ ឬបរាជ័យរបស់បុគ្គលជាក់លាក់ណាមួយក្នុងការរស់រានមានជីវិត និងការចាកចេញពីកូនចៅ។ ភាវៈមានជីវិតទាំងអស់មានសក្ដានុពលក្នុងការផលិតចំនួនដ៏ច្រើននៃប្រភេទរបស់វា។ ជាឧទាហរណ៍ កូនចៅដែល daphnia មួយ (សត្វក្រៀលទឹកសាប) អាចទុកនៅរដូវក្ដៅឈានដល់ទំហំតារាសាស្ត្រ - ច្រើនជាង 10 30 បុគ្គល ដែលលើសពីម៉ាស់ផែនដី។
ទោះជាយ៉ាងណាក៏ដោយ ការលូតលាស់ដែលមិនអាចកាត់ថ្លៃបាននៃចំនួនសារពាង្គកាយមានជីវិត គឺមិនត្រូវបានគេសង្កេតឃើញជាក់ស្តែងនោះទេ។ តើអ្វីជាហេតុផលសម្រាប់បាតុភូតនេះ? បុគ្គលភាគច្រើនបានស្លាប់នៅដំណាក់កាលផ្សេងៗនៃការអភិវឌ្ឍន៍ ហើយមិនបន្សល់ទុកកូនចៅទេ។ មានហេតុផលជាច្រើនដែលកំណត់ការរីកលូតលាស់នៃចំនួនសត្វ៖ ទាំងនេះគឺជាកត្តាធម្មជាតិ និងអាកាសធាតុ និងការប្រយុទ្ធប្រឆាំងនឹងបុគ្គលនៃប្រភេទសត្វដទៃទៀត និងប្រភេទរបស់វាផ្ទាល់។
វាត្រូវបានគេដឹងថាអត្រានៃការបន្តពូជរបស់បុគ្គលនៃប្រភេទណាមួយកាន់តែខ្ពស់ អត្រាមរណភាពកាន់តែធ្ងន់ធ្ងរ។ ជាឧទាហរណ៍ Beluga ពងប្រហែលមួយលានពងក្នុងអំឡុងពេលពង ហើយមានតែផ្នែកតូចមួយនៃសាច់ដែលឈានដល់វ័យពេញវ័យ។ រុក្ខជាតិក៏ផលិតគ្រាប់ពូជក្នុងបរិមាណដ៏ច្រើនផងដែរ ប៉ុន្តែនៅក្រោមលក្ខខណ្ឌធម្មជាតិ មានតែផ្នែកតូចមួយនៃពួកវាប៉ុណ្ណោះដែលផ្តល់កំណើតដល់រុក្ខជាតិថ្មី។ ភាពខុសគ្នារវាងលទ្ធភាពនៃប្រភេទសត្វសម្រាប់ការបន្តពូជគ្មានដែនកំណត់ និងធនធានមានកំណត់គឺជាហេតុផលចម្បងសម្រាប់ការតស៊ូដើម្បីអត្ថិភាព។ ការស្លាប់របស់កូនចៅកើតឡើងដោយសារហេតុផលផ្សេងៗ។ វាអាចជាជម្រើស ឬចៃដន្យ (ការស្លាប់របស់បុគ្គលនៅក្នុងភ្លើងឆេះព្រៃ ក្នុងករណីទឹកជំនន់ ការអន្តរាគមន៍របស់មនុស្សក្នុងធម្មជាតិ។ល។)។
ការតស៊ូមិនជាក់លាក់។អាំងតង់ស៊ីតេនៃការបន្តពូជ និងការស្លាប់ដោយជ្រើសរើសរបស់បុគ្គលដែលសម្របខ្លួនមិនបានល្អទៅនឹងការផ្លាស់ប្តូរលក្ខខណ្ឌបរិស្ថានមានសារៈសំខាន់ជាទីបំផុតសម្រាប់ការផ្លាស់ប្តូរការវិវត្តន៍។ មិនគួរគិតថា បុគ្គលដែលមានលក្ខណៈមិនគួរគប្បីប្រាកដជាស្លាប់ឡើយ ។ មានប្រូបាប៊ីលីតេខ្ពស់ដែលនាងនឹងបន្សល់ទុកកូនចៅតិចជាង ឬគ្មានទាល់តែសោះ ចំណែកបុគ្គលធម្មតានឹងបន្តពូជ។ ដូច្នេះហើយ អ្នកដែលរស់រានមានជីវិត និងបន្តពូជ ជាក្បួនគឺសមជាង។ នេះគឺជាយន្តការចម្បងនៃការជ្រើសរើសធម្មជាតិ។ ការស្លាប់ដោយជ្រើសរើសរបស់មនុស្សមួយចំនួន និងការរស់រានមានជីវិតរបស់បុគ្គលផ្សេងទៀតគឺជាបាតុភូតដែលជាប់ទាក់ទងគ្នាដែលមិនអាចកាត់ថ្លៃបាន។
វាគឺនៅក្នុងសេចក្តីថ្លែងការណ៍ដ៏សាមញ្ញ និងនៅ glance ដំបូងនេះដែលថាទេពកោសល្យនៃគំនិតរបស់ Darwin នៃការជ្រើសរើសធម្មជាតិគឺ ពោលគឺការបន្តពូជនៃបុគ្គលដែលសមជាងដែលឈ្នះការតស៊ូដើម្បីអត្ថិភាព។ ការតស៊ូរបស់បុគ្គលនៅក្នុងប្រភេទមួយគឺមានលក្ខណៈចម្រុះបំផុត។
នេះប្រហែលជាការតស៊ូផ្ទាល់សម្រាប់អត្ថិភាព (ការប្រកួតប្រជែង) រវាងបុគ្គលដែលមានប្រភេទដូចគ្នាសម្រាប់ប្រភពអាហារ ទឹក ទីជម្រក ទឹកដីសំបុក។ល។ បុគ្គលនៃប្រភេទសត្វដូចគ្នាប្រកួតប្រជែងគ្នាទៅវិញទៅមកក្នុងលក្ខខណ្ឌនៃភាពធន់ទ្រាំទៅនឹងកត្តាបរិស្ថាន biotic និង abiotic មិនអំណោយផល: ជំងឺឆ្លង សត្វមំសាសី សីតុណ្ហភាពខ្លាំង។ល។
ទំនាក់ទំនងរវាងបុគ្គលនៅក្នុងប្រភេទមួយមិនត្រូវបានកំណត់ចំពោះការតស៊ូ និងការប្រកួតប្រជែងនោះទេ វាក៏មានជំនួយទៅវិញទៅមកផងដែរ។
ជំនួយទៅវិញទៅមកត្រូវបានបង្ហាញយ៉ាងច្បាស់បំផុតនៅក្នុងគ្រួសារ និងក្រុមនៃអង្គការសត្វ នៅពេលដែលបុគ្គលខ្លាំង និងធំការពារកូនឆ្មា និងញី ការពារទឹកដី និងសត្វព្រៃ រួមចំណែកដល់ភាពជោគជ័យរបស់ក្រុមទាំងមូល ឬគ្រួសារទាំងមូល ជារឿយៗត្រូវចំណាយប្រាក់ ជីវិតរបស់ពួកគេ។
ជំនួយទៅវិញទៅមករវាងបុគ្គលដែលជាកម្មសិទ្ធិរបស់ក្រុមគ្រួសារតែមួយ ហើយដូច្នេះ ការមានហ្សែនទូទៅមិនកាត់បន្ថយភាពធ្ងន់ធ្ងរនៃការតស៊ូដើម្បីមានជីវិតនោះទេ ប៉ុន្តែផ្ទេរវាទៅយន្តហោះផ្សេង។ ការប្រកួតប្រជែងរវាងបុគ្គលត្រូវបានជំនួសដោយការប្រកួតប្រជែងរវាងក្រុមពាក់ព័ន្ធ។ ជំនួយទៅវិញទៅមកខ្លួនវាក្លាយជាឧបករណ៍នៃការតស៊ូដើម្បីអត្ថិភាព។ ប៉ុន្តែលទ្ធផលនៅតែដដែល - ពីមួយជំនាន់ទៅមួយជំនាន់ក្នុងចំនួនប្រជាជន ភាពញឹកញាប់នៃហ្សែនដែលផ្តល់នូវកាយសម្បទាខ្ពស់ក្នុងន័យទូលំទូលាយនៃពាក្យកើនឡើង។
ម៉ាស៊ីនសំខាន់នៃការផ្លាស់ប្តូរការវិវត្តន៍គឺការជ្រើសរើសធម្មជាតិនៃសារពាង្គកាយដែលសម្របខ្លួនបំផុតដែលកើតឡើងជាលទ្ធផលនៃការតស៊ូដើម្បីអត្ថិភាព។
Interspecies តស៊ូ។ការតស៊ូអន្តរជាក់លាក់គួរតែត្រូវបានយល់ថាជាទំនាក់ទំនងរវាងបុគ្គល ប្រភេទផ្សេងគ្នា. ពួកគេអាចប្រកួតប្រជែងបាន ដោយផ្អែកលើផលប្រយោជន៍ទៅវិញទៅមក ឬអព្យាក្រឹត។ ការប្រកួតប្រជែងអន្តរជាក់លាក់ឈានដល់ភាពធ្ងន់ធ្ងរជាពិសេសនៅក្នុងករណីដែលប្រភេទសត្វដែលរស់នៅក្នុងលក្ខខណ្ឌអេកូឡូស៊ីស្រដៀងគ្នា និងប្រើប្រាស់ប្រភពអាហារដូចគ្នាប្រកួតប្រជែង។ ជាលទ្ធផលនៃការតស៊ូអន្តរជាក់លាក់ ទាំងការផ្លាស់ទីលំនៅនៃប្រភេទសត្វប្រឆាំងមួយកើតឡើង ឬការសម្របខ្លួននៃប្រភេទសត្វទៅនឹងលក្ខខណ្ឌផ្សេងៗនៅក្នុងតំបន់តែមួយ ឬទីបំផុតការបំបែកទឹកដីរបស់ពួកគេ។
ប្រភេទថ្មពីរប្រភេទអាចបង្ហាញពីផលវិបាកនៃការតស៊ូរវាងប្រភេទសត្វដែលទាក់ទងគ្នាយ៉ាងជិតស្និទ្ធ។ នៅកន្លែងទាំងនោះដែលជួរនៃប្រភេទសត្វទាំងនេះត្រួតលើគ្នា ពោលគឺបក្សីនៃប្រភេទទាំងពីររស់នៅក្នុងទឹកដីតែមួយ ប្រវែងនៃចំពុះ និងវិធីដែលពួកគេទទួលបានអាហារមានភាពខុសគ្នាខ្លាំង។ នៅក្នុងតំបន់ជម្រកដែលមិនត្រួតស៊ីគ្នានៃ nuthatches មិនមានភាពខុសប្លែកគ្នានៅក្នុងប្រវែងចំពុះ និងវិធីសាស្រ្តនៃការទទួលបានអាហារទេ។ ការតស៊ូអន្តរជាក់លាក់ នាំទៅដល់ការបំបែកផ្នែកអេកូឡូស៊ី និងភូមិសាស្រ្តនៃប្រភេទសត្វ។
ប្រសិទ្ធភាពនៃការជ្រើសរើស។ប្រសិទ្ធភាពនៃការជ្រើសរើសធម្មជាតិអាស្រ័យទៅលើអាំងតង់ស៊ីតេរបស់វា និងស្តុកនៃការប្រែប្រួលតំណពូជដែលបានបង្គរនៅក្នុងចំនួនប្រជាជន។ អាំងតង់ស៊ីតេនៃការជ្រើសរើសត្រូវបានកំណត់ដោយសមាមាត្រនៃបុគ្គលណាដែលរស់បានដល់ភាពចាស់ទុំខាងផ្លូវភេទ និងចូលរួមក្នុងការបន្តពូជ។ សមាមាត្រនេះកាន់តែតូច អាំងតង់ស៊ីតេនៃការជ្រើសរើសកាន់តែច្រើន។ ទោះជាយ៉ាងណាក៏ដោយ សូម្បីតែការជ្រើសរើសដែលពឹងផ្អែកខ្លាំងបំផុតក៏នឹងគ្មានប្រសិទ្ធភាពដែរ ប្រសិនបើការប្រែប្រួលនៃបុគ្គលនៅក្នុងចំនួនប្រជាជនមិនសំខាន់ ឬវាមានលក្ខណៈមិនតំណពូជ។ សម្រាប់ការជ្រើសរើសដើម្បីផ្លាស់ប្តូរតម្លៃមធ្យមនៃលក្ខណៈ បុគ្គលនៅក្នុងចំនួនប្រជាជនត្រូវតែមានហ្សែនខុសគ្នាពីគ្នាទៅវិញទៅមក។ នេះត្រូវបានបញ្ជាក់ដោយអ្នកជំនាញហ្សែនជនជាតិដាណឺម៉ាក V. Johansen ជាមួយនឹងការពិសោធន៍បុរាណរបស់គាត់។ គាត់ដាច់ឆ្ងាយពីសណ្តែក បន្ទាត់ស្អាតដែលត្រូវបានទទួលដោយការ pollination ដោយខ្លួនឯងនៃរុក្ខជាតិដើមមួយ និងកូនចៅរបស់វានៅក្នុងស៊េរីនៃជំនាន់មួយ។ បន្ទាត់ដែលបានបង្កើតតាមរបៀបនេះគឺមានលក្ខណៈដូចគ្នាសម្រាប់ហ្សែនភាគច្រើន ពោលគឺនៅក្នុងជួរ ភាពប្រែប្រួលគឺគ្រាន់តែជាលក្ខណៈនៃការកែប្រែប៉ុណ្ណោះ។ នៅក្នុងបន្ទាត់បែបនេះ ការជ្រើសរើសទំហំសណ្តែកមិនបាននាំទៅដល់ការរីកធំ ឬការថយចុះនៃជំនាន់ជាបន្តបន្ទាប់នោះទេ។ នៅក្នុងពពួកសណ្តែកធម្មតា ភាពប្រែប្រួលនៃតំណពូជមាន ហើយការជ្រើសរើសបានប្រែទៅជាមានប្រសិទ្ធភាព។
- តើអ្វីទៅជាហេតុផលនៃការតស៊ូដើម្បីអត្ថិភាព?
- តើការតស៊ូដើម្បីអត្ថិភាពឈប់នៅក្នុងហ្វូងសត្វទាំងនោះដែលមានជំនួយទៅវិញទៅមកឬ?
- តើវាអាចនាំឱ្យមានផលវិបាកអ្វីខ្លះ? ការតស៊ូអន្តរជាក់លាក់?
- តើអ្វីកំណត់ប្រសិទ្ធភាពនៃការជ្រើសរើសធម្មជាតិ?
ការជ្រើសរើសធម្មជាតិគឺជាដំណើរការវិវត្តន៍ដ៏សំខាន់ ជាលទ្ធផលនៃចំនួនបុគ្គលដែលមានកាយសម្បទាអតិបរមា (លក្ខណៈអំណោយផលបំផុត) ក្នុងចំនួនប្រជាជនកើនឡើង ខណៈដែលចំនួនបុគ្គលដែលមានលក្ខណៈមិនអំណោយផលមានការថយចុះ។
ការជ្រើសរើសធម្មជាតិគឺជាកត្តាដឹកនាំក្នុងដំណើរការវិវត្តន៍ ដែលជាកម្លាំងជំរុញនៃការវិវត្តន៍។
ទិសដៅនៃការជ្រើសរើសធម្មជាតិត្រូវបានគេហៅថាវ៉ិចទ័រជ្រើសរើស។
មានវិធីសាស្រ្តជាច្រើនដើម្បីកំណត់គំនិតនៃ "ការជ្រើសរើសធម្មជាតិ" ។
តាមទស្សនៈនៃទ្រឹស្តីសំយោគបុរាណនៃការវិវត្តន៍៖
ការជ្រើសរើសធម្មជាតិគឺជាសំណុំនៃដំណើរការជីវសាស្រ្តដែលធានាដល់ការបន្តពូជខុសៗគ្នានៃព័ត៌មានហ្សែននៅក្នុងចំនួនប្រជាជន។
លទ្ធផលនៃការជ្រើសរើសធម្មជាតិ៖
1. ការអភិរក្សរចនាសម្ព័ន្ធហ្សែនរបស់ប្រជាជន
2. ការផ្លាស់ប្តូររចនាសម្ព័ន្ធហ្សែនរបស់ប្រជាជន
3. ការលេចឡើងនៃវ៉ារ្យ៉ង់ថ្មីនៃលក្ខណៈដែលមានពីមុន
4. ការលេចចេញនូវមុខងារថ្មីៗជាមូលដ្ឋាន
5. ការបង្កើតប្រភេទសត្វថ្មី។
6. ធម្មជាតិរីកចម្រើននៃការវិវត្តន៍ជីវសាស្រ្ត។
នៅក្នុងដំណើរការនៃការជ្រើសរើសធម្មជាតិការផ្លាស់ប្តូរត្រូវបានជួសជុលដែលបង្កើនភាពរឹងមាំនៃសារពាង្គកាយ។ ការជ្រើសរើសធម្មជាតិត្រូវបានគេហៅថាយន្តការ "បង្ហាញឱ្យឃើញដោយខ្លួនឯង" ព្រោះវាធ្វើតាមការពិតសាមញ្ញដូចជា:
1. សារពាង្គកាយបង្កើតកូនចៅច្រើនជាងអាចរស់បាន;
2. មានមរតកនៅក្នុងចំនួនប្រជាជននៃសារពាង្គកាយទាំងនេះ។ ភាពអាចផ្លាស់ប្តូរបាន;
3. សារពាង្គកាយដែលមានលក្ខណៈហ្សែនខុសៗគ្នា មានអត្រារស់រានមានជីវិតខុសៗគ្នា និងសមត្ថភាពក្នុងការបន្តពូជ។
លក្ខខណ្ឌបែបនេះបង្កើតការប្រកួតប្រជែងរវាងសារពាង្គកាយសម្រាប់ការរស់រានមានជីវិត និងការបន្តពូជ ហើយជាលក្ខខណ្ឌចាំបាច់អប្បបរមាសម្រាប់ការវិវត្តន៍តាមរយៈការជ្រើសរើសធម្មជាតិ។ ដូច្នេះហើយ សារពាង្គកាយដែលមានលក្ខណៈតំណពូជដែលផ្តល់ឱ្យពួកគេនូវគុណសម្បត្តិប្រកួតប្រជែង ទំនងជានឹងបញ្ជូនពួកវាទៅឱ្យកូនចៅរបស់ពួកគេ ជាងសារពាង្គកាយដែលមានលក្ខណៈតំណពូជដែលមិនមានអត្ថប្រយោជន៍បែបនេះ។
គោលគំនិតកណ្តាលនៃគំនិតនៃការជ្រើសរើសធម្មជាតិគឺការសម្របខ្លួននៃសារពាង្គកាយ។ សម្បទាត្រូវបានកំណត់ថាជាសមត្ថភាពរបស់សារពាង្គកាយមួយក្នុងការរស់រានមានជីវិត និងបន្តពូជ ដែលកំណត់ទំហំនៃការចូលរួមចំណែកហ្សែនរបស់វាចំពោះមនុស្សជំនាន់ក្រោយ។ ទោះយ៉ាងណាក៏ដោយ រឿងសំខាន់ក្នុងការកំណត់កាយសម្បទាមិនមែនជាចំនួនកូនចៅសរុបទេ ប៉ុន្តែចំនួនកូនចៅដែលមានហ្សែនដែលបានផ្តល់ឱ្យ ( សម្បទាដែលទាក់ទង) ជាឧទាហរណ៍ ប្រសិនបើកូនចៅរបស់សារពាង្គកាយដែលទទួលបានជោគជ័យ និងបន្តពូជយ៉ាងឆាប់រហ័សខ្សោយ ហើយមិនអាចបន្តពូជបានល្អ នោះការរួមចំណែកហ្សែន ហើយដូច្នេះ ភាពរឹងមាំនៃសារពាង្គកាយនោះនឹងមានកម្រិតទាប។
28
. យន្តការនៃភាពឯកោអន្តរជាក់លាក់
គំនិតនៃប្រភេទជីវសាស្រ្តសន្មតថាអត្ថិភាពនៃភាពឯកោនៃការបន្តពូជដែលមានលក្ខណៈជាក់លាក់ - ពោលគឺភាពឯកោដែលរារាំងបុគ្គលដែលជាកម្មសិទ្ធិរបស់ប្រភេទផ្សេងៗគ្នាពីការបង្កាត់ពូជ។ ភាពឯកោបន្តពូជធានាមិនត្រឹមតែការរួមរស់នៃប្រភេទសត្វដែលទាក់ទងគ្នាយ៉ាងជិតស្និទ្ធប៉ុណ្ណោះទេ ប៉ុន្តែក៏មានឯករាជ្យភាពនៃការវិវត្តរបស់ពួកគេផងដែរ។
ភាពខុសគ្នាមួយត្រូវបានធ្វើឡើងរវាងអ៊ីសូឡង់បឋម និងអនុវិទ្យាល័យ។ ភាពឯកោបឋមកើតឡើងដោយគ្មានការចូលរួមពីការជ្រើសរើសធម្មជាតិ; ទម្រង់នៃភាពឯកោនេះគឺចៃដន្យ និងមិនអាចទាយទុកជាមុនបាន។ ភាពឯកោបន្ទាប់បន្សំកើតឡើងក្រោមឥទ្ធិពលនៃភាពស្មុគស្មាញនៃកត្តាវិវត្តន៍បឋម។ ទម្រង់នៃភាពឯកោនេះកើតឡើងដោយធម្មជាតិ និងអាចព្យាករណ៍បាន។
ទម្រង់សាមញ្ញបំផុតនៃភាពឯកោអន្តរជាក់លាក់គឺ លំហ, ឬ ភូមិសាស្ត្រអ៊ីសូឡង់។ ប្រភេទសត្វមិនអាចបង្កាត់ពូជបានទេ ពីព្រោះចំនួនប្រជាជននៃប្រភេទផ្សេងៗគ្នាគឺដាច់ឆ្ងាយពីគ្នាទៅវិញទៅមក។ ដោយផ្អែកលើកម្រិតនៃភាពឯកោនៃលំហ ប្រជាជន allopatric, adjacent-sympatric និង biotic-sympatric populations ត្រូវបានសម្គាល់។
ចំនួនប្រជាជនស៊ីមេទ្រីជីវសាស្រ្តអាចបង្កាត់គ្នាទៅវិញទៅមកដើម្បីបង្កើតជាកូនកាត់អន្តរជាក់លាក់។ ប៉ុន្តែបន្ទាប់មក ដោយសារតែការបង្កើតកូនកាត់ជាបន្តបន្ទាប់ និងការឆ្លងកាត់របស់វាជាមួយនឹងទម្រង់មាតាបិតា នោះប្រភេទសត្វសុទ្ធត្រូវតែរលាយបាត់ភ្លាមៗ។ ទោះជាយ៉ាងណាក៏ដោយ ការពិតវាមិនកើតឡើងទេ ដែលបង្ហាញពីអត្ថិភាពនៃយន្តការជាច្រើនដែលការពារយ៉ាងមានប្រសិទ្ធភាពក្នុងការបង្កាត់អន្តរជាក់លាក់នៅក្នុង លក្ខខណ្ឌធម្មជាតិដែលត្រូវបានបង្កើតឡើងដោយមានការចូលរួមនៃទម្រង់ជាក់លាក់នៃការជ្រើសរើសធម្មជាតិ ដែលគេស្គាល់ថាជា "ដំណើរការ Wallace"។ (នេះជាមូលហេតុដែលការឆ្លងកាត់អេកូឡូស៊ី-ភូមិសាស្ត្ររវាងប្រភេទសត្វដែលមិនទាក់ទងគ្នាក្រោមលក្ខខណ្ឌធម្មជាតិគឺជោគជ័យបំផុត)។
ជាធម្មតា យន្តការឯកោចំនួនបីក្រុមត្រូវបានសម្គាល់៖ precopulatory, prezygotic និង postzygotic ។ ទន្ទឹមនឹងនេះយន្តការឯកោ prezygotic និង postzygotic ត្រូវបានរួមបញ្ចូលគ្នាជាញឹកញាប់ក្រោមឈ្មោះទូទៅ "យន្តការ postcopulatory" ។
មានដាន។ យន្តការនៃភាពឯកោនៃការបន្តពូជ interspecific: 1. យន្តការ precopulatory - ការពារការ copulation (មិត្តរួមនៅក្នុងសត្វឬការ pollination នៅក្នុងរុក្ខជាតិ) ។ ក្នុងករណីនេះ ទាំង gametes មាតាបិតា ឬមាតា (និងហ្សែនដែលត្រូវគ្នា) ត្រូវបានលុបចោល។ 2. យន្តការ Prezygotic - ការពារការបង្កកំណើត។ ក្នុងករណីនេះ gametes មាតា (ហ្សែន) ត្រូវបានលុបចោល ប៉ុន្តែ gametes មាតា (ហ្សែន) ត្រូវបានរក្សា។ ភាពឯកោ Prezygotic អាចជាបឋម ឬអនុវិទ្យាល័យ។ 3. យន្តការ Postzygotic - ការពារការបញ្ជូនហ្សែនពីប្រភេទមាតាបិតាទៅជំនាន់បន្តបន្ទាប់តាមរយៈកូនកាត់។
29
. ភាពចម្រុះជីវសាស្រ្ត។ កម្រិតនៃជីវចម្រុះ intraspecific
ភាពចម្រុះនៃជីវសាស្រ្ត - អត្ថិភាពនៃប្រភេទរុក្ខជាតិ និងសត្វជាច្រើនប្រភេទ - គឺជាលក្ខខណ្ឌដែលមិនអាចខ្វះបានសម្រាប់ការរស់រានមានជីវិតរបស់មនុស្ស។ អនុសញ្ញាអង្គការសហប្រជាជាតិស្តីពីភាពចម្រុះជីវសាស្រ្ត (1992) ដែលប្រទេសចំនួន 190 បានយល់ព្រម មានគោលបំណងការពារ និងអភិរក្សប្រភេទសត្វ និងរុក្ខជាតិចម្រុះ និងជម្រករបស់ពួកគេ។ អនុសញ្ញានេះមានកាតព្វកិច្ចការពារជីវចម្រុះ និងធានាវា។ ការអភិវឌ្ឍប្រកបដោយចីរភាពនិងផ្តល់នូវការចែករំលែកដោយយុត្តិធម៌ និងសមធម៌នៃអត្ថប្រយោជន៍ដែលកើតចេញពីការប្រើប្រាស់ធនធានហ្សែន។ ពិធីការ Cartagena របស់នាង
ដែលចូលជាធរមានក្នុងឆ្នាំ 2003 និងមានគោលបំណងធានានូវការប្រើប្រាស់ប្រកបដោយសុវត្ថិភាពនៃសារពាង្គកាយកែប្រែហ្សែន បច្ចុប្បន្នត្រូវបានចុះហត្ថលេខាដោយប្រទេសចំនួន 143 ។ ភាពចម្រុះនៃជីវសាស្រ្ត សំដៅលើទាំងអស់នៃ "សារពាង្គកាយមានជីវិតខុសៗគ្នា ភាពប្រែប្រួលក្នុងចំនោមពួកវា និងស្មុគស្មាញអេកូឡូស៊ីដែលពួកវាជាផ្នែក ដែលរួមមានភាពចម្រុះក្នុងប្រភេទសត្វ រវាងប្រភេទសត្វ និងប្រព័ន្ធអេកូឡូស៊ី"។ ក្នុងករណីនេះ ចាំបាច់ត្រូវបែងចែករវាងភាពចម្រុះសកល និងក្នុងស្រុក។ ភាពចម្រុះជីវសាស្រ្ត គឺជាធនធានជីវសាស្រ្តដ៏សំខាន់បំផុតមួយ ( ធនធានជីវសាស្រ្តត្រូវបានចាត់ទុកថាជា "សម្ភារៈហ្សែន សារពាង្គកាយ ឬផ្នែករបស់វា ឬប្រព័ន្ធអេកូឡូស៊ីដែលបានប្រើ ឬមានសក្តានុពលមានប្រយោជន៍សម្រាប់មនុស្សជាតិ រួមទាំងតុល្យភាពធម្មជាតិនៅក្នុង និងរវាងប្រព័ន្ធអេកូឡូស៊ី")។
ប្រភេទនៃភាពចម្រុះជីវសាស្រ្តខាងក្រោមត្រូវបានសម្គាល់៖ អាល់ហ្វា បេតា ហ្គាម៉ា និងភាពចម្រុះហ្សែន។ ដោយ α-ភាពចម្រុះ យើងមានន័យថា ភាពចម្រុះនៃប្រភេទ ដោយ β-ភាពចម្រុះ យើងមានន័យថា ភាពចម្រុះនៃសហគមន៍នៅលើ ទឹកដីជាក់លាក់; γ-ភាពចម្រុះគឺជាសូចនាករសំខាន់ដែលរួមបញ្ចូល α- និង β-ភាពចម្រុះ។ ទោះជាយ៉ាងណាក៏ដោយ មូលដ្ឋាននៃប្រភេទជីវចម្រុះដែលបានរាយបញ្ជីគឺ ភាពចម្រុះនៃហ្សែន (អន្តរជាក់លាក់ ភាពចម្រុះ)។
វត្តមាននៃអាឡែលពីរឬច្រើន (ហើយតាមប្រភេទហ្សែន) ក្នុងចំនួនប្រជាជនត្រូវបានគេហៅថា polymorphism ហ្សែន. វាត្រូវបានទទួលយកជាធម្មតាថាប្រេកង់នៃ allele កម្របំផុតនៅក្នុង polymorphism គួរតែមានយ៉ាងហោចណាស់ 1% (0.01) ។ អត្ថិភាពនៃហ្សែនហ្សែនគឺជាតម្រូវការជាមុនសម្រាប់ការអភិរក្សជីវចម្រុះ។
គំនិតអំពីតំរូវការដើម្បីរក្សាហ្សែនហ្សែននៅក្នុងប្រជាជនធម្មជាតិត្រូវបានបង្កើតឡើងនៅទសវត្សរ៍ឆ្នាំ 1920 ។ ជនរួមជាតិដ៏អស្ចារ្យរបស់យើង។ Nikolai Ivanovich Vavilov បានបង្កើតគោលលទ្ធិនៃសម្ភារៈប្រភពនិងបញ្ជាក់ពីតម្រូវការក្នុងការបង្កើតឃ្លាំងនៃហ្សែនពិភពលោកនៃរុក្ខជាតិដាំដុះ។ Alexander Sergeevich Serebrovsky បានបង្កើតគោលលទ្ធិនៃហ្សែនហ្សែន។ គោលគំនិតនៃ "ហ្សែនហ្សែន" រួមបញ្ចូលភាពចម្រុះនៃហ្សែននៃប្រភេទសត្វដែលបានអភិវឌ្ឍកំឡុងពេលការវិវត្តន៍ ឬការជ្រើសរើសរបស់វា ហើយផ្តល់នូវសមត្ថភាពក្នុងការសម្របខ្លួន និងផលិតរបស់វា។ លោក Sergei Sergeevich Chetverikov បានដាក់មូលដ្ឋានគ្រឹះនៃគោលលទ្ធិ និងវិធីសាស្រ្តសម្រាប់វាយតម្លៃពីភាពតំណពូជហ្សែននៃចំនួនប្រជាជននៃប្រភេទសត្វព្រៃ និងរុក្ខជាតិ។
30. បញ្ហានៃការអភិរក្សពពួកពពួកពពួក polymorphism នៅដំណាក់កាលផ្សេងៗនៃ speciation
ការជួសជុលដោយចៃដន្យនៃ alleles អព្យាក្រឹតជ្រើសរើសដ៏កម្រគឺអាចធ្វើទៅបានជាលទ្ធផលនៃតំណពូជហ្សែនតែនៅក្នុងចំនួនប្រជាជនតិចតួចប៉ុណ្ណោះ។ ប៉ុន្តែនៅក្នុងចំនួនប្រជាជនបែបនេះ អាឡែសអព្យាក្រឹតជ្រើសរើសនៃហ្សែនផ្សេងទៀតក៏ត្រូវបានកត់ត្រាដោយចៃដន្យផងដែរ ដែលគួរតែកាត់បន្ថយកម្រិតនៃពហុហ្សែនហ្សែន។ វាត្រូវបានបង្កើតឡើងដែលផ្ទាំងទឹកកកបានជះឥទ្ធិពលគួរឱ្យកត់សម្គាល់លើរចនាសម្ព័ន្ធចំនួនប្រជាជននៃប្រភេទត្រីមួយចំនួន ឧទាហរណ៍ ត្រីសាម៉ុងប៉ាស៊ីហ្វិក។ ក្នុងករណីភាគច្រើន ចំនួនប្រជាជននៃប្រភេទសត្វទំនើបត្រូវបានកំណត់លក្ខណៈដោយកម្រិតខ្ពស់នៃពហុមុខងារហ្សែន។ យន្តការពិតនៃការបង្កើតភាពឯកោក្រោយ copulatory គឺស្មុគស្មាញជាងអ្វីដែលបានពិភាក្សាខាងលើ។
ដោយផ្អែកលើកម្រិតនៃភាពចម្រុះ intraspecific ក្រុមខ្លាំងពីរនៃប្រភេទអាចត្រូវបានសម្គាល់: ជាមួយនឹងកម្រិតខ្ពស់និងទាបនៃ polymorphism intraspecific ។ ក្រុមទី 1 គឺជាប្រភេទ polytypic eurybiont ដែលមានជួរធំទូលាយ និងរចនាសម្ព័ន្ធ intraspecific ស្មុគស្មាញ ជាមួយនឹងកម្រិតខ្ពស់នៃ intrapopulation និង interpopulation variability ។ ក្រុមទី 2 គឺជាក្រុមដែលមានកម្រិតទាបនៃភាពប្រែប្រួល។ វាច្បាស់ណាស់ថាក្រុមទីមួយនៃប្រភេទសត្វមានសក្តានុពលវិវត្តន៍ខ្ពស់ពោលគឺឧ។ អាចផ្តល់ការកើនឡើងដល់ប្រភេទសត្វថ្មីជាច្រើន (ហើយជាបន្តបន្ទាប់ទៅពន្ធនៃឋានៈខ្ពស់ជាង)។ ក្រុមទីពីរត្រូវបានកំណត់លក្ខណៈដោយសក្តានុពលវិវត្តន៍ទាប; លទ្ធភាពដែលវានឹងផ្តល់ការកើនឡើងដល់ប្រភេទសត្វថ្មី (និងជាពិសេសពន្ធដារដែលមានឋានៈខ្ពស់ជាង) គឺតិចជាងច្រើន។
31. វឌ្ឍនភាពជីវសាស្រ្ត និងលក្ខណៈវិនិច្ឆ័យរបស់វា។ ស្ថេរភាពជីវសាស្រ្ត។ តំរែតំរង់ជីវសាស្រ្តនិងមូលហេតុរបស់វា។
វឌ្ឍនភាពជីវសាស្រ្តកំណត់ក្រុមបុគ្គលនៃសារពាង្គកាយនៅដំណាក់កាលជាក់លាក់នៃការអភិវឌ្ឍន៍ ពិភពសរីរាង្គ.
លក្ខណៈវិនិច្ឆ័យសម្រាប់វឌ្ឍនភាពជីវសាស្រ្ត៖
1. បង្កើនចំនួនបុគ្គលនៃក្រុមដែលកំពុងពិចារណា។
2. ការពង្រីកតំបន់។
3. ទម្រង់ដែលពឹងផ្អែកខ្លាំង និងជាក់លាក់។
ជាលទ្ធផលវាត្រូវបានគេសង្កេតឃើញ ចូលទៅក្នុងតំបន់អាដាប់ធ័រថ្មីជាមួយនឹងវិទ្យុសកម្មបន្សាំជាបន្តបន្ទាប់ពោលគឺការចែកចាយនៅតាមទីជម្រកផ្សេងៗ។ បច្ចុប្បន្ននេះ angiosperms សត្វល្អិត សត្វស្លាប និងថនិកសត្វ គឺស្ថិតនៅក្នុងស្ថានភាពនៃដំណើរការជីវសាស្រ្ត។
មានវិធីសំខាន់ៗចំនួនបីដើម្បីសម្រេចបាននូវវឌ្ឍនភាពជីវសាស្រ្ត៖ arogenesis, allogenesis និង catagenesis ដែលជំនួសគ្នាទៅវិញទៅមកដោយធម្មជាតិ។
Arogenesis- ដំណើរការផ្សព្វផ្សាយ កម្រិតទូទៅអង្គការ.
លក្ខណៈវិនិច្ឆ័យសម្រាប់ arogenesis (វឌ្ឍនភាព morphophysiological):
ក) ជាប្រព័ន្ធ- ការកែលម្អប្រព័ន្ធ homeostasis និង homeoresis;
ខ) ថាមពល- បង្កើនប្រសិទ្ធភាព សារពាង្គកាយក្នុងករណីជាក់លាក់មួយ - ការបង្កើនកម្រិតនៃការរំលាយអាហារ (បក្សីថនិកសត្វ);
វី) ព័ត៌មាន- ការកើនឡើងនៃបរិមាណព័ត៌មាន៖ ហ្សែន (ការកើនឡើងនៃបរិមាណ DNA នៅក្នុងកោសិកា) និង epigenetic (ការចងចាំ ការរៀនសូត្រ) ។
លទ្ធផលនៃវឌ្ឍនភាពគឺ វឌ្ឍនភាពជីវសាស្រ្តទូទៅ ពាក់ព័ន្ធនឹងការចូលទៅក្នុងតំបន់បន្សាំថ្មី។
Aromorphoses មានទំហំធំ ការសម្របខ្លួនជាមុនដែលផ្តល់ឱ្យសារពាង្គកាយនូវឱកាសដើម្បីរស់នៅក្នុងលក្ខខណ្ឌថ្មីជាមុន។ ជាលទ្ធផលនៃ aromorphoses, ធំទូលាយមួយ។ វិទ្យុសកម្មអាដាប់ធ័រ. វិទ្យុសកម្មអាដាប់ធ័រគឺជាការបំបែកនៃដើមបុព្វបុរសនៃក្រុមនៃសារពាង្គកាយទៅជាសាខាដាច់ដោយឡែកពីគ្នាក្នុងដំណើរនៃការវិវត្តន៍បន្សាំ។
Allogenesis គឺជាដំណើរការនៃរូបរាងនៃការបន្សាំឯកជននៅក្នុងលក្ខខណ្ឌរស់នៅមួយចំនួន មិនត្រូវបានអមដោយការកើនឡើងនៃកម្រិតទូទៅនៃអង្គការនោះទេ។ ជាលទ្ធផលនៃ allogenesis, allomorphoses, telomorphoses និង hypermorphoses ត្រូវបានបង្កើតឡើង។
Allomorphoses គឺជាការសម្របខ្លួនតាមកាយវិភាគសាស្ត្រ និង morphological ដែលធានាឱ្យមានការសម្របខ្លួនទៅនឹងលក្ខខណ្ឌរស់នៅមួយចំនួន។
Telomorphoses ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការផ្លាស់ប្តូរពី បរិស្ថានទូទៅឯកជន មានកំណត់។
Hypermorphoses គឺជាសញ្ញា hypertrophied ។ ឧទាហរណ៍មួយគឺភាពមហិមា។
សេចក្តីផ្តើម
1. Charles Darwin - ស្ថាបនិកទ្រឹស្តីនៃការវិវត្តន៍
2. មូលហេតុ និងទម្រង់នៃ "ការតស៊ូដើម្បីអត្ថិភាព" នៅក្នុងធម្មជាតិរស់នៅ
3. ទ្រឹស្តីនៃការជ្រើសរើសធម្មជាតិ ទម្រង់នៃការជ្រើសរើសធម្មជាតិ
4. តួនាទីនៃការប្រែប្រួលតំណពូជក្នុងការវិវត្តន៍នៃប្រភេទសត្វ
សេចក្តីសន្និដ្ឋាន
ការណែនាំ
ពាក្យ "ការវិវត្តន៍" (មកពីឡាតាំងវិវឌ្ឍន៍ - ការដាក់ពង្រាយ) ត្រូវបានប្រើជាលើកដំបូងនៅក្នុងស្នាដៃអំប្រ៊ីយ៉ុងដោយអ្នកជំនាញធម្មជាតិជនជាតិស្វីស Charles Bonnet ក្នុងឆ្នាំ 1762។ បច្ចុប្បន្ននេះ ការវិវត្តន៍ត្រូវបានគេយល់ថាជាដំណើរការដែលមិនអាចត្រឡប់វិញនៃការផ្លាស់ប្តូរប្រព័ន្ធដែលកើតឡើងតាមពេលវេលា។ អ្វីដែលកើតឡើងថ្មី ខុសប្លែកពីគេ ឈរនៅដំណាក់កាលនៃការអភិវឌ្ឍខ្ពស់ជាង។
ដំណើរការនៃការវិវត្តន៍ទាក់ទងនឹងបាតុភូតជាច្រើនដែលកើតឡើងនៅក្នុងធម្មជាតិ។ ឧទាហរណ៍ តារាវិទូនិយាយអំពីការវិវត្តនៃប្រព័ន្ធភព និងផ្កាយ អ្នកភូគព្ភវិទូ - អំពីការវិវត្តនៃផែនដី ជីវវិទូ - អំពីការវិវត្តនៃសត្វមានជីវិត។ ទន្ទឹមនឹងនេះពាក្យ "ការវិវត្តន៍" ជារឿយៗត្រូវបានអនុវត្តចំពោះបាតុភូតដែលមិនទាក់ទងដោយផ្ទាល់ទៅនឹងធម្មជាតិក្នុងន័យតូចចង្អៀតនៃពាក្យ។ ឧទាហរណ៍ ពួកគេនិយាយអំពីការវិវត្តនៃប្រព័ន្ធសង្គម ទស្សនៈ ម៉ាស៊ីន ឬសម្ភារៈមួយចំនួន។ល។
គោលគំនិតនៃការវិវត្តន៍ត្រូវផ្អែកលើអត្ថន័យពិសេសនៅក្នុងវិទ្យាសាស្ត្រធម្មជាតិ ដែលការវិវត្តន៍ជីវសាស្រ្តត្រូវបានសិក្សាជាចម្បង។ ការវិវត្តន៍ជីវសាស្រ្តគឺជាការមិនអាចត្រឡប់វិញបាន និងក្នុងកម្រិតជាក់លាក់មួយដែលបានដឹកនាំការអភិវឌ្ឍជាប្រវត្តិសាស្ត្រនៃធម្មជាតិរស់នៅ អមដោយការផ្លាស់ប្តូរនៅក្នុងសមាសភាពហ្សែននៃចំនួនប្រជាជន ការបង្កើតការសម្របខ្លួន ការបង្កើត និងការផុតពូជនៃប្រភេទសត្វ ការផ្លាស់ប្តូរនៃ biogeocenoses និង biosphere ទាំងមូល។ ម្យ៉ាងវិញទៀត ការវិវត្តន៍ជីវសាស្រ្តគួរត្រូវបានយល់ថាជាដំណើរការនៃការអភិវឌ្ឍន៍ប្រវត្តិសាស្រ្តសម្របខ្លួននៃទម្រង់រស់នៅនៅគ្រប់កម្រិតនៃការរៀបចំរបស់ភាវៈរស់។
ទ្រឹស្ដីនៃការវិវត្តន៍ត្រូវបានបង្កើតឡើងដោយ Charles Darwin (1809-1882) ហើយបានរៀបរាប់នៅក្នុងសៀវភៅរបស់គាត់ដែលមានចំណងជើងថា The Origin of Species by Means of Natural Selection ឬ ការអភិរក្សពូជដែលពេញចិត្តក្នុងការតស៊ូដើម្បីជីវិត (1859)។
1. C. Darwin - ស្ថាបនិកនៃទ្រឹស្តីនៃការវិវត្តន៍
Charles Darwin កើតនៅថ្ងៃទី 12 ខែកុម្ភៈ ឆ្នាំ 1809។ នៅក្នុងគ្រួសាររបស់វេជ្ជបណ្ឌិត។ ពេលកំពុងសិក្សានៅសាកលវិទ្យាល័យ Edinburgh និង Cambridge លោក Darwin បានទទួលចំណេះដឹងជ្រៅជ្រះអំពីសត្វវិទ្យា រុក្ខសាស្ត្រ និងភូគព្ភសាស្ត្រ និងជំនាញ និងរសជាតិសម្រាប់ការស្រាវជ្រាវវាល។
សៀវភៅរបស់អ្នកភូគព្ភវិទូអង់គ្លេសដ៏ឆ្នើម Charles Lyell "គោលការណ៍ភូគព្ភសាស្ត្រ" បានដើរតួនាទីយ៉ាងសំខាន់ក្នុងការបង្កើតទស្សនៈពិភពលោកបែបវិទ្យាសាស្ត្ររបស់គាត់។ Lyell បានប្រកែកដូច្នេះ រូបរាងទំនើបផែនដីត្រូវបានបង្កើតឡើងបន្តិចម្តងៗ ក្រោមឥទ្ធិពលនៃកម្លាំងធម្មជាតិដដែល ដែលនៅតែសកម្មរហូតមកដល់សព្វថ្ងៃនេះ។ ដាវីនបានស្គាល់គំនិតវិវត្តន៍របស់ Erasmus Darwin, Lamarck និងអ្នកវិវត្តន៍ដំបូងផ្សេងទៀត ប៉ុន្តែគាត់មិនបានរកឃើញថាពួកគេគួរឱ្យជឿនោះទេ។
វេនសម្រេចចិត្តនៅក្នុងជោគវាសនារបស់គាត់គឺការធ្វើដំណើររបស់គាត់ជុំវិញពិភពលោកនៅលើកប៉ាល់ Beagle (1832-1837) ។ ការសង្កេតដែលបានធ្វើឡើងក្នុងអំឡុងពេលធ្វើដំណើរនេះបានបម្រើជាមូលដ្ឋានគ្រឹះសម្រាប់ទ្រឹស្តីនៃការវិវត្តន៍។ យោងតាមលោក Darwin ខ្លួនគាត់ផ្ទាល់ ក្នុងអំឡុងពេលធ្វើដំណើរនេះ គាត់មានការចាប់អារម្មណ៍ខ្លាំងបំផុតដោយ៖ “១) ការរកឃើញហ្វូស៊ីលសត្វយក្ស ដែលត្រូវបានគ្របដោយសំបកស្រដៀងនឹងសំបករបស់ armadillos ទំនើប។ 2) ការពិតដែលថានៅពេលដែលយើងផ្លាស់ទីឆ្លងកាត់ទ្វីបអាមេរិកខាងត្បូង ប្រភេទសត្វដែលទាក់ទងយ៉ាងជិតស្និទ្ធជំនួសគ្នាទៅវិញទៅមក។ 3) ការពិតដែលថាប្រភេទសត្វដែលទាក់ទងយ៉ាងជិតស្និទ្ធនៃកោះផ្សេងៗនៃប្រជុំកោះ Galapagos ខុសគ្នាបន្តិចបន្តួចពីគ្នាទៅវិញទៅមក។ វាច្បាស់ណាស់ថាប្រភេទនៃការពិតទាំងនេះក៏ដូចជារឿងជាច្រើនទៀតអាចពន្យល់បានតែលើមូលដ្ឋាននៃការសន្មត់ថាប្រភេទសត្វកំពុងផ្លាស់ប្តូរបន្តិចម្តងៗ ហើយបញ្ហានេះបានចាប់ផ្តើមលងខ្ញុំ។
ពេលត្រឡប់ពីការធ្វើដំណើររបស់គាត់ ដាវីនចាប់ផ្តើមគិតអំពីបញ្ហានៃប្រភពដើមនៃប្រភេទសត្វ។ គាត់ពិចារណាលើគំនិតផ្សេងៗ រួមទាំងគំនិតរបស់ Lamarck ហើយបដិសេធពួកគេ ដោយសារគ្មាននរណាម្នាក់ក្នុងចំណោមពួកគេពន្យល់ពីការពិតនៃភាពប្រែប្រួលដ៏អស្ចារ្យនៃសត្វ និងរុក្ខជាតិចំពោះជីវភាពរស់នៅរបស់ពួកគេ។ អ្វីដែលអ្នកវិវត្តន៍ដំបូងបានគិតថាជាការផ្តល់ឱ្យ ហើយការពន្យល់ដោយខ្លួនឯងហាក់ដូចជាសំណួរសំខាន់បំផុតសម្រាប់ដាវីន។ វាប្រមូលទិន្នន័យអំពីការប្រែប្រួលនៃសត្វ និងរុក្ខជាតិនៅក្នុងធម្មជាតិ និងក្រោមការចិញ្ចឹម។ ជាច្រើនឆ្នាំក្រោយមក ដោយនឹកចាំពីរបៀបដែលទ្រឹស្ដីរបស់គាត់កើតឡើង ដាវីននឹងសរសេរថា៖ «ខ្ញុំដឹងភ្លាមៗថា មូលដ្ឋានគ្រឹះនៃភាពជោគជ័យរបស់មនុស្សក្នុងការបង្កើតពូជសត្វ និងរុក្ខជាតិមានប្រយោជន៍គឺការជ្រើសរើស។ ទោះយ៉ាងណាក៏ដោយ សម្រាប់ពេលខ្លះ វានៅតែជាអាថ៌កំបាំងសម្រាប់ខ្ញុំពីរបៀបដែលការជ្រើសរើសអាចត្រូវបានអនុវត្តចំពោះសារពាង្គកាយដែលរស់នៅក្រោមលក្ខខណ្ឌធម្មជាតិ»។ នៅពេលនោះ គំនិតរបស់អ្នកវិទ្យាសាស្ត្រអង់គ្លេស T. Malthus អំពីការបង្កើនចំនួនប្រជាជនក្នុងដំណើរការធរណីមាត្រត្រូវបានពិភាក្សាយ៉ាងខ្លាំងក្លានៅក្នុងប្រទេសអង់គ្លេស។ លោក Darwin បន្តថា៖ «ក្នុងខែតុលា ឆ្នាំ 1838 ខ្ញុំបានអានសៀវភៅ Malthus's On Population ហើយដោយសារការសង្កេតយ៉ាងយូរអំពីរបៀបរស់នៅរបស់សត្វ និងរុក្ខជាតិ ខ្ញុំបានត្រៀមខ្លួនយ៉ាងល្អដើម្បីដឹងគុណចំពោះសារៈសំខាន់នៃការតស៊ូជាសកលសម្រាប់អត្ថិភាពរបស់ខ្ញុំ។ ភ្លាមៗនោះត្រូវបានវាយប្រហារដោយគំនិតថានៅក្រោមលក្ខខណ្ឌបែបនេះ ការផ្លាស់ប្តូរអំណោយផលគួរមាននិន្នាការបន្ត ហើយអ្វីដែលមិនអំណោយផលនឹងត្រូវបំផ្លាញចោល។ លទ្ធផលនៃការនេះគួរតែជាការបង្កើតប្រភេទសត្វថ្មី»។
ដូច្នេះគំនិតនៃប្រភពដើមនៃប្រភេទសត្វតាមរយៈការជ្រើសរើសធម្មជាតិបានកើតចេញពីដាវីនក្នុងឆ្នាំ 1838 ។ គាត់បានធ្វើការលើវាអស់រយៈពេល 20 ឆ្នាំ។ នៅឆ្នាំ 1856 តាមដំបូន្មានរបស់ Lyell គាត់បានចាប់ផ្តើមរៀបចំការងាររបស់គាត់សម្រាប់ការបោះពុម្ពផ្សាយ។ នៅឆ្នាំ 1858 អ្នកវិទ្យាសាស្ត្រអង់គ្លេសវ័យក្មេង Alfred Wallace បានផ្ញើសាត្រាស្លឹករឹតរបស់ Darwin នៃអត្ថបទរបស់គាត់ "On the tendency of Varieties to deviate Unlimitedly from the Original Type"។ អត្ថបទនេះមានការបង្ហាញអំពីគំនិតនៃប្រភពដើមនៃប្រភេទសត្វតាមរយៈការជ្រើសរើសធម្មជាតិ។ ដាវីនបានត្រៀមខ្លួនរួចជាស្រេចក្នុងការបដិសេធមិនបោះពុម្ពស្នាដៃរបស់គាត់ ប៉ុន្តែមិត្តភក្តិរបស់គាត់គឺលោក Charles Lyell ភូគព្ភវិទូ និងរុក្ខសាស្ត្រ G. Hooker ដែលបានស្គាល់ជាយូរមកហើយអំពីគំនិតរបស់ដាវីន និងស្គាល់សេចក្តីព្រាងបឋមនៃសៀវភៅរបស់គាត់បានបញ្ចុះបញ្ចូលអ្នកវិទ្យាសាស្ត្រថាការងារទាំងពីរគួរតែត្រូវបានបោះពុម្ពក្នុងពេលដំណាលគ្នា។ .
សៀវភៅរបស់ Darwin, On the Origin of Species by Means of Natural Selection, or the Preservation of Favored Races in the Struggle for Life ត្រូវបានបោះពុម្ពនៅឆ្នាំ 1859 ហើយភាពជោគជ័យរបស់វាលើសពីការរំពឹងទុកទាំងអស់។ គំនិតនៃការវិវត្តន៍របស់គាត់បានជួបជាមួយនឹងការគាំទ្រយ៉ាងខ្លាំងពីអ្នកវិទ្យាសាស្ត្រមួយចំនួន និងការរិះគន់យ៉ាងចាស់ដៃពីអ្នកដទៃ។ ស្នាដៃបន្តបន្ទាប់របស់ដាវីន "ការផ្លាស់ប្តូរសត្វ និងរុក្ខជាតិអំឡុងពេលរស់នៅក្នុងផ្ទះ" "តំណពូជរបស់មនុស្ស និងការជ្រើសរើសផ្លូវភេទ" និង "ការបង្ហាញពីអារម្មណ៍នៅក្នុងមនុស្ស និងសត្វ" ត្រូវបានបកប្រែភ្លាមៗទៅជាភាសាជាច្រើនបន្ទាប់ពីការបោះពុម្ពផ្សាយរបស់ពួកគេ។ . គួរកត់សម្គាល់ថាការបកប្រែជាភាសារុស្សីនៃសៀវភៅរបស់ដាវីនដែលមានចំណងជើងថា "ការផ្លាស់ប្តូរសត្វនិងរុក្ខជាតិនៅក្រោមផ្ទះ" ត្រូវបានបោះពុម្ពមុនជាងអត្ថបទដើមរបស់វា។ បុរាណវិទូជនជាតិរុស្សីដ៏ឆ្នើម V. O. Kovalevsky បានបកប្រែសៀវភៅនេះពីភ័ស្តុតាងដែលផ្តល់ឱ្យគាត់ដោយដាវីនហើយបោះពុម្ពវានៅក្នុងបញ្ហាដាច់ដោយឡែក។
ទ្រឹស្ដីវិវត្តន៍របស់ដាវីន គឺជាគោលលទ្ធិរួមនៃការអភិវឌ្ឍន៍ប្រវត្តិសាស្ត្រនៃពិភពសរីរាង្គ។ វាគ្របដណ្តប់លើបញ្ហាជាច្រើន ដែលសំខាន់បំផុតនោះគឺភស្តុតាងនៃការវិវត្តន៍ ការកំណត់អត្តសញ្ញាណកម្លាំងជំរុញនៃការវិវត្តន៍ ការកំណត់ផ្លូវ និងលំនាំនៃដំណើរការវិវត្តន៍។ល។
ខ្លឹមសារនៃការបង្រៀនវិវត្តន៍ស្ថិតនៅក្នុងគោលការណ៍មូលដ្ឋានដូចខាងក្រោម៖
1. សត្វមានជីវិតគ្រប់ប្រភេទដែលរស់នៅលើផែនដី មិនដែលបង្កើតដោយនរណាម្នាក់ឡើយ។
2. ដោយបានបង្កើតឡើងតាមធម្មជាតិ ទម្រង់សរីរាង្គត្រូវបានផ្លាស់ប្តូរបន្តិចម្តងៗ និងប្រសើរឡើង ស្របតាមលក្ខខណ្ឌបរិស្ថាន។
3. ការផ្លាស់ប្តូរនៃប្រភេទសត្វនៅក្នុងធម្មជាតិគឺផ្អែកលើលក្ខណៈសម្បត្តិនៃសារពាង្គកាយដូចជាតំណពូជ និងភាពប្រែប្រួល ក៏ដូចជាការជ្រើសរើសធម្មជាតិដែលកើតឡើងជានិច្ចនៅក្នុងធម្មជាតិ។ ការជ្រើសរើសធម្មជាតិកើតឡើងតាមរយៈអន្តរកម្មស្មុគស្មាញនៃសារពាង្គកាយជាមួយគ្នា និងដោយកត្តា ធម្មជាតិគ្មានជីវិត; ដាវីនបានហៅទំនាក់ទំនងនេះថាជាការតស៊ូដើម្បីអត្ថិភាព។
4. លទ្ធផលនៃការវិវត្តន៍គឺការសម្របខ្លួនរបស់សារពាង្គកាយទៅនឹងលក្ខខណ្ឌរស់នៅរបស់ពួកគេ និងភាពសម្បូរបែបនៃប្រភេទសត្វនៅក្នុងធម្មជាតិ។
2. ហេតុផល និងទម្រង់នៃ "ការតស៊ូដើម្បីអត្ថិភាព"
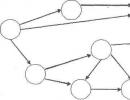
"ការតស៊ូដើម្បីអត្ថិភាព" គឺជាគំនិតមួយដែល Charles Darwin ប្រើដើម្បីកំណត់លក្ខណៈនៃទំនាក់ទំនងទាំងមូលរវាងបុគ្គល និងកត្តាបរិស្ថានផ្សេងៗ។ ទំនាក់ទំនងទាំងនេះកំណត់ភាពជោគជ័យ ឬបរាជ័យរបស់បុគ្គលជាក់លាក់ណាមួយក្នុងការរស់រានមានជីវិត និងការចាកចេញពីកូនចៅ។ ភាវៈមានជីវិតទាំងអស់មានសក្ដានុពលក្នុងការផលិតចំនួនដ៏ច្រើននៃប្រភេទរបស់វា។ ជាឧទាហរណ៍ កូនចៅដែល daphnia មួយ (សត្វក្រៀលទឹកសាប) អាចទុកនៅរដូវក្ដៅ ឈានដល់ទំហំតារាសាស្ត្រ ច្រើនជាង 10 30 បុគ្គល ដែលលើសពីម៉ាស់ផែនដី។ ទោះជាយ៉ាងណាក៏ដោយ ការលូតលាស់ដែលមិនអាចកាត់ថ្លៃបាននៃចំនួនសារពាង្គកាយមានជីវិត គឺមិនត្រូវបានគេសង្កេតឃើញជាក់ស្តែងនោះទេ។ តើអ្វីជាហេតុផលសម្រាប់បាតុភូតនេះ? បុគ្គលភាគច្រើនបានស្លាប់នៅដំណាក់កាលផ្សេងៗនៃការអភិវឌ្ឍន៍ ហើយមិនបន្សល់ទុកកូនចៅទេ។ មានហេតុផលជាច្រើនដែលកំណត់ការលូតលាស់នៃចំនួនសត្វ៖ ទាំងនេះគឺជាកត្តាធម្មជាតិ និងអាកាសធាតុ និងការប្រយុទ្ធប្រឆាំងនឹងបុគ្គលនៃប្រភេទសត្វរបស់ពួកគេ និងប្រភេទដទៃទៀត។
រូបភាពទី 1 - សកម្មភាពនៃការតស៊ូដើម្បីអត្ថិភាព
វាត្រូវបានគេដឹងថាអត្រានៃការបន្តពូជរបស់បុគ្គលនៃប្រភេទណាមួយកាន់តែខ្ពស់ ការស្លាប់កាន់តែធ្ងន់ធ្ងរ។ ជាឧទាហរណ៍ Beluga ពងប្រហែលមួយលានពងក្នុងអំឡុងពេលពង ហើយមានតែផ្នែកតូចមួយនៃពួកវាប៉ុណ្ណោះដែលឈានដល់ការលូតលាស់ពេញវ័យ។ រុក្ខជាតិក៏ផលិតគ្រាប់ពូជក្នុងបរិមាណដ៏ច្រើនផងដែរ ប៉ុន្តែនៅក្រោមលក្ខខណ្ឌធម្មជាតិ មានតែផ្នែកតូចមួយនៃពួកវាប៉ុណ្ណោះដែលផ្តល់កំណើតដល់រុក្ខជាតិថ្មី។ ភាពខុសគ្នារវាងលទ្ធភាពនៃប្រភេទសត្វសម្រាប់ការបន្តពូជគ្មានដែនកំណត់ និងធនធានមានកំណត់គឺជាហេតុផលចម្បងសម្រាប់ការតស៊ូដើម្បីអត្ថិភាព។ ការស្លាប់របស់កូនចៅកើតឡើងដោយសារហេតុផលផ្សេងៗ។ វាអាចជាជម្រើស ឬចៃដន្យ (ក្នុងករណីទឹកជំនន់ អន្តរាគមន៍របស់មនុស្សក្នុងធម្មជាតិ ភ្លើងឆេះព្រៃ។ល។)។

រូបភាពទី 2 - ទម្រង់នៃការតស៊ូដើម្បីអត្ថិភាព
ការតស៊ូមិនជាក់លាក់។អាំងតង់ស៊ីតេនៃការបន្តពូជ និងការស្លាប់ដោយជ្រើសរើសរបស់បុគ្គលដែលសម្របខ្លួនមិនបានល្អទៅនឹងការផ្លាស់ប្តូរលក្ខខណ្ឌបរិស្ថានមានសារៈសំខាន់ជាទីបំផុតសម្រាប់ការផ្លាស់ប្តូរការវិវត្តន៍។ មិនគួរគិតថា បុគ្គលដែលមានលក្ខណៈមិនគួរគប្បីប្រាកដជាស្លាប់ឡើយ ។ មានប្រូបាប៊ីលីតេខ្ពស់ដែលនាងនឹងបន្សល់ទុកកូនចៅតិចជាង ឬគ្មានទាល់តែសោះ ចំណែកបុគ្គលធម្មតានឹងបន្តពូជ។ អាស្រ័យហេតុនេះ សត្វដែលសាកសមបំផុតតែងតែរស់រានមានជីវិត និងបន្តពូជ។ នេះគឺជាយន្តការចម្បងនៃការជ្រើសរើសធម្មជាតិ។ ការស្លាប់ដោយជ្រើសរើសរបស់មនុស្សមួយចំនួន និងការរស់រានមានជីវិតរបស់បុគ្គលផ្សេងទៀតគឺជាបាតុភូតដែលជាប់ទាក់ទងគ្នាដែលមិនអាចកាត់ថ្លៃបាន។ វាគឺនៅក្នុងសេចក្តីថ្លែងការណ៍ដ៏សាមញ្ញមួយ និងនៅ glance ដំបូងដែលថា ទេពកោសល្យនៃគំនិតរបស់ Darwin នៃការជ្រើសរើសធម្មជាតិគឺ ពោលគឺឧ។ ក្នុងការបន្តពូជនៃបុគ្គលដែលសម្របខ្លួនកាន់តែច្រើនដែលឈ្នះការតស៊ូដើម្បីអត្ថិភាព។ ការតស៊ូរបស់បុគ្គលនៅក្នុងប្រភេទមួយគឺមានលក្ខណៈចម្រុះបំផុត។
បុគ្គលមិនត្រឹមតែប្រកួតប្រជែងដើម្បីប្រភពអាហារ សំណើម ព្រះអាទិត្យ និងទឹកដីប៉ុណ្ណោះទេ ប៉ុន្តែជួនកាលចូលរួមក្នុងការប្រយុទ្ធដោយផ្ទាល់។
នៅក្នុងសត្វ dioecious បុរសនិងញីមានភាពខុសគ្នាជាចម្បងនៅក្នុងរចនាសម្ព័ន្ធនៃសរីរាង្គបន្តពូជរបស់ពួកគេ។ ទោះជាយ៉ាងណាក៏ដោយ ភាពខុសគ្នាច្រើនតែពង្រីកដល់សញ្ញាខាងក្រៅ និងអាកប្បកិរិយា។ ចងចាំសម្លៀកបំពាក់ដ៏ភ្លឺស្វាងរបស់សត្វមាន់ សិតសក់ធំ ដុះលើជើងរបស់គាត់ និងការច្រៀងដ៏ធំ។ សត្វស្លាបឈ្មោលគឺស្អាតណាស់បើធៀបនឹងសត្វមាន់ដែលល្មមៗជាង។ សត្វឆ្កែនៃថ្គាមខាងលើ - ភ្លុក - លូតលាស់យ៉ាងខ្លាំងជាពិសេសនៅក្នុង walrus បុរស។ ភាពខុសគ្នាខាងក្រៅនៅក្នុងរចនាសម្ព័ន្ធនៃភេទត្រូវបានគេហៅថា dimorphism ផ្លូវភេទ ហើយដោយសារតែតួនាទីរបស់ពួកគេក្នុងការជ្រើសរើសផ្លូវភេទ។ ការជ្រើសរើសផ្លូវភេទ គឺជាការប្រកួតប្រជែងរវាងបុរស ដើម្បីមានឱកាសបន្តពូជ។ គោលបំណងនេះត្រូវបានបម្រើដោយការច្រៀង ការបង្ហាញអាកប្បកិរិយា ភាពស្និទ្ធស្នាល និងការឈ្លោះប្រកែកគ្នាជាញឹកញាប់រវាងបុរស។
ភាពចម្រុះផ្លូវភេទ និងការជ្រើសរើសផ្លូវភេទគឺរីករាលដាលយ៉ាងខ្លាំងនៅក្នុងពិភពសត្វ រួមទាំងសត្វព្រូនផងដែរ។ ទម្រង់នៃការជ្រើសរើសនេះគួរតែត្រូវបានចាត់ទុកថាជាករណីពិសេសនៃការជ្រើសរើសធម្មជាតិដែលមិនជាក់លាក់។
ទំនាក់ទំនងរបស់បុគ្គលនៅក្នុងប្រភេទសត្វមិនត្រូវបានកំណត់ចំពោះការតស៊ូ និងការប្រកួតប្រជែងនោះទេ។ ក៏មានជំនួយទៅវិញទៅមក។ ជំនួយទៅវិញទៅមករបស់បុគ្គល ការកំណត់ទឹកដីបុគ្គល - ទាំងអស់នេះកាត់បន្ថយភាពធ្ងន់ធ្ងរនៃអន្តរកម្មអន្តរកម្ម។
ជំនួយទៅវិញទៅមកត្រូវបានបង្ហាញយ៉ាងច្បាស់បំផុតនៅក្នុងអង្គការគ្រួសារ និងក្រុមសត្វ។ នៅពេលដែលបុគ្គលខ្លាំង និងធំការពារកូន និងញី ការពារទឹកដី និងសត្វព្រៃ រួមចំណែកដល់ភាពជោគជ័យរបស់ក្រុមទាំងមូល ឬគ្រួសារទាំងមូល ជារឿយៗត្រូវចំណាយអស់មួយជីវិត។ ការបន្តពូជ និងការស្លាប់របស់បុគ្គលម្នាក់ៗទទួលបានលក្ខណៈជ្រើសរើសតាមរយៈការប្រកួតប្រជែងរវាងបុគ្គលចម្រុះហ្សែនក្នុងចំនួនប្រជាជនដែលបានផ្តល់ឱ្យ ដូច្នេះការតស៊ូផ្ទៃក្នុងគឺជាហេតុផលដ៏សំខាន់បំផុតសម្រាប់ការជ្រើសរើសធម្មជាតិ។ ម៉ាស៊ីនសំខាន់នៃការផ្លាស់ប្តូរការវិវត្តន៍គឺការជ្រើសរើសធម្មជាតិនៃសារពាង្គកាយដែលសម្របខ្លួនបំផុតដែលកើតឡើងជាលទ្ធផលនៃការតស៊ូដើម្បីអត្ថិភាព។
Interspecies ប្រយុទ្ធ . ការតស៊ូអន្តរជាក់លាក់គួរតែត្រូវបានយល់ថាជាការតស៊ូរបស់បុគ្គលដែលមានប្រភេទផ្សេងៗគ្នា។ ការតស៊ូរវាងប្រភេទសត្វឈានដល់ភាពធ្ងន់ធ្ងរជាពិសេសនៅក្នុងករណីដែលប្រភេទសត្វដែលរស់នៅក្នុងលក្ខខណ្ឌអេកូឡូស៊ីស្រដៀងគ្នា និងប្រើប្រាស់ប្រភពអាហារដូចគ្នាប្រកួតប្រជែង។ ជាលទ្ធផលនៃការតស៊ូអន្តរជាក់លាក់ ទាំងការផ្លាស់ទីលំនៅនៃប្រភេទសត្វប្រឆាំងមួយកើតឡើង ឬការផ្លាស់ទីលំនៅរបស់ប្រភេទសត្វទៅលក្ខខណ្ឌផ្សេងៗគ្នានៅក្នុងតំបន់តែមួយ ឬចុងក្រោយគឺការបំបែកទឹកដីរបស់ពួកគេ។
ប្រភេទថ្មពីរប្រភេទអាចបង្ហាញពីផលវិបាកនៃការតស៊ូរវាងប្រភេទសត្វដែលទាក់ទងគ្នាយ៉ាងជិតស្និទ្ធ។ នៅក្នុងកន្លែងដែលជួរនៃប្រភេទសត្វទាំងនេះត្រួតលើគ្នា, i.e. សត្វស្លាបទាំងពីរប្រភេទរស់នៅលើទ្រឹស្ដីដូចគ្នា ប្រវែងនៃចំពុះ និងលទ្ធភាពទទួលបានអាហារមានភាពខុសគ្នាខ្លាំង។ នៅក្នុងតំបន់ជម្រកដែលមិនត្រួតស៊ីគ្នានៃ nuthatches មិនមានភាពខុសប្លែកគ្នានៅក្នុងប្រវែងចំពុះ និងវិធីសាស្រ្តនៃការទទួលបានអាហារទេ។ ការតស៊ូអន្តរជាក់លាក់ នាំទៅដល់ការបំបែកផ្នែកអេកូឡូស៊ី និងភូមិសាស្រ្តនៃប្រភេទសត្វ។
3. ការប្រយុទ្ធប្រឆាំងនឹងលក្ខខណ្ឌមិនអំណោយផលនៃធម្មជាតិអសរីរាង្គក៏បង្កើនការប្រកួតប្រជែងមិនជាក់លាក់ផងដែរ ដោយសារបុគ្គលនៃប្រភេទដូចគ្នាប្រកួតប្រជែងសម្រាប់អាហារ ពន្លឺ ភាពកក់ក្តៅ និងលក្ខខណ្ឌផ្សេងទៀតនៃអត្ថិភាព។ វាមិនមែនជារឿងចៃដន្យទេដែលរុក្ខជាតិមួយនៅក្នុងវាលខ្សាច់ត្រូវបានគេនិយាយថាដើម្បីប្រឆាំងនឹងគ្រោះរាំងស្ងួត។ នៅក្នុង tundra ដើមឈើត្រូវបានតំណាងដោយទម្រង់មនុស្សតឿទោះបីជាពួកគេមិនមានបទពិសោធន៍ប្រកួតប្រជែងពីរុក្ខជាតិផ្សេងទៀតក៏ដោយ។ អ្នកឈ្នះក្នុងការប្រយុទ្ធគឺជាបុគ្គលដែលអាចសម្រេចបានបំផុត (ដំណើរការសរីរវិទ្យា និងការរំលាយអាហាររបស់ពួកគេដំណើរការកាន់តែមានប្រសិទ្ធភាព)។ ប្រសិនបើលក្ខណៈជីវសាស្រ្តត្រូវបានទទួលមរតក នោះនៅទីបំផុតនឹងនាំទៅដល់ការកែលម្អការសម្របតាមប្រភេទសត្វទៅនឹងបរិស្ថាន។
3. ទ្រឹស្តីនៃការជ្រើសរើសធម្មជាតិ
ទម្រង់នៃការជ្រើសរើសធម្មជាតិ
ការជ្រើសរើសកើតឡើងជាបន្តបន្ទាប់ជាបន្តបន្ទាប់ជាបន្តបន្ទាប់ជាបន្តបន្ទាប់ និងរក្សាជាចម្បងទម្រង់ទាំងនោះដែលស៊ីគ្នាជាងនឹងលក្ខខណ្ឌដែលបានផ្តល់ឱ្យ។ ការជ្រើសរើសតាមធម្មជាតិ និងការលុបបំបាត់បុគ្គលមួយចំនួននៃប្រភេទសត្វមួយត្រូវបានផ្សារភ្ជាប់គ្នាដោយ inextricably និងជាលក្ខខណ្ឌចាំបាច់សម្រាប់ការវិវត្តនៃប្រភេទសត្វនៅក្នុងធម្មជាតិ។
គ្រោងការណ៍នៃសកម្មភាពនៃការជ្រើសរើសធម្មជាតិនៅក្នុងប្រព័ន្ធប្រភេទមួយយោងទៅតាម Darwin មានដូចខាងក្រោម៖
1) ការបំរែបំរួលគឺជាលក្ខណៈនៃក្រុមសត្វ និងរុក្ខជាតិណាមួយ ហើយសារពាង្គកាយមានភាពខុសគ្នាពីគ្នាទៅវិញទៅមកក្នុងការគោរពជាច្រើន។
2) ចំនួនសារពាង្គកាយនៃប្រភេទសត្វនីមួយៗដែលកើតលើសពីចំនួនសត្វដែលអាចស្វែងរកអាហារ និងរស់បាន។ ទោះជាយ៉ាងណាក៏ដោយចាប់តាំងពីចំនួននៃប្រភេទនីមួយៗគឺថេរនៅក្រោមលក្ខខណ្ឌធម្មជាតិវាគួរតែត្រូវបានសន្មត់ថា ភាគច្រើនកូនចៅស្លាប់។ ប្រសិនបើកូនចៅទាំងអស់នៃប្រភេទសត្វណាមួយនៅរស់រានមានជីវិត និងបន្តពូជ ពួកគេនឹងជំនួសប្រភេទសត្វផ្សេងទៀតទាំងអស់នៅលើផែនដីឆាប់ៗនេះ។
៣) ដោយសារបុគ្គលកើតមកច្រើនជាងអាចរស់បាន នោះមានការតស៊ូដើម្បីអត្ថិភាព ការប្រកួតប្រជែងអាហារ និងទីជម្រក។ នេះអាចជាការតស៊ូយ៉ាងសកម្មក្នុងជីវិត និងសេចក្តីស្លាប់ ឬមិនសូវច្បាស់ ប៉ុន្តែមិនមានការប្រកួតប្រជែងដ៏មានប្រសិទ្ធភាពនោះទេ ដូចជាឧទាហរណ៍ សម្រាប់រុក្ខជាតិក្នុងអំឡុងពេលគ្រោះរាំងស្ងួត ឬត្រជាក់។
4) ក្នុងចំណោមការផ្លាស់ប្តូរជាច្រើនដែលបានសង្កេតឃើញនៅក្នុងសត្វមានជីវិត ខ្លះជួយសម្រួលដល់ការរស់រានមានជីវិតក្នុងការតស៊ូដើម្បីអត្ថិភាព ចំណែកខ្លះទៀតនាំទៅដល់ការស្លាប់របស់ម្ចាស់របស់វា។ គំនិតនៃ "ការរស់រានមានជីវិតនៃសមបំផុត" គឺជាស្នូលនៃទ្រឹស្តីនៃការជ្រើសរើសធម្មជាតិ;
5) បុគ្គលដែលរស់រានមានជីវិតផ្តល់ភាពរីកចម្រើនដល់មនុស្សជំនាន់ក្រោយ ហើយដូច្នេះការផ្លាស់ប្តូរ "ជោគជ័យ" ត្រូវបានបញ្ជូនទៅមនុស្សជំនាន់ក្រោយៗទៀត។ ជាលទ្ធផល ជំនាន់បន្តបន្ទាប់នីមួយៗ ប្រែជាសម្របខ្លួនទៅនឹងបរិយាកាសរបស់វាកាន់តែច្រើន។ នៅពេលដែលបរិស្ថានផ្លាស់ប្តូរ ការសម្របខ្លួនបន្ថែមទៀតកើតឡើង។ ប្រសិនបើការជ្រើសរើសតាមធម្មជាតិដំណើរការអស់ជាច្រើនឆ្នាំ នោះពូជចុងក្រោយអាចប្រែទៅជាខុសគ្នាខ្លាំងពីបុព្វបុរសរបស់ពួកគេ ដែលវាត្រូវបានគេណែនាំឱ្យបំបែកពួកវាទៅជាប្រភេទឯករាជ្យ។
វាក៏អាចកើតឡើងផងដែរដែលថាសមាជិកមួយចំនួននៃក្រុមបុគ្គលណាមួយទទួលបានការផ្លាស់ប្តូរជាក់លាក់ ហើយរកឃើញថាពួកគេសម្របតាម បរិស្ថាននៅក្នុងវិធីមួយ ខណៈពេលដែលសមាជិកផ្សេងទៀតរបស់ខ្លួនដែលមានសំណុំផ្សេងគ្នានៃការផ្លាស់ប្តូរនឹងត្រូវបានសម្របតាមវិធីផ្សេង។ តាមវិធីនេះ ពីប្រភេទដូនតាមួយ បានផ្តល់ថាក្រុមស្រដៀងគ្នានេះនៅដាច់ពីគេ ពីរ ឬច្រើនប្រភេទអាចកើតឡើង។
ការជ្រើសរើសការបើកបរ។ការជ្រើសរើសតាមធម្មជាតិតែងតែនាំឱ្យមានការកើនឡើងនៃសម្បទាជាមធ្យមរបស់ប្រជាជន។ ការផ្លាស់ប្តូរនៅក្នុងលក្ខខណ្ឌខាងក្រៅអាចនាំឱ្យមានការផ្លាស់ប្តូរនៅក្នុងកាយសម្បទានៃ genotypes បុគ្គល។ ដើម្បីឆ្លើយតបទៅនឹងការផ្លាស់ប្តូរទាំងនេះ ការជ្រើសរើសធម្មជាតិ ដោយប្រើអាងស្តុកទឹកដ៏ធំនៃភាពចម្រុះនៃហ្សែននៅទូទាំងជាច្រើន។ សញ្ញាផ្សេងគ្នានាំឱ្យមានការផ្លាស់ប្តូរយ៉ាងសំខាន់នៅក្នុងរចនាសម្ព័ន្ធហ្សែននៃប្រជាជន។ ប្រសិនបើបរិយាកាសខាងក្រៅកំពុងផ្លាស់ប្តូរជានិច្ចក្នុងទិសដៅជាក់លាក់មួយ នោះការជ្រើសរើសធម្មជាតិផ្លាស់ប្តូររចនាសម្ព័ន្ធហ្សែនរបស់ប្រជាជនតាមរបៀបដែលសម្បទារបស់វានៅក្នុងលក្ខខណ្ឌផ្លាស់ប្តូរទាំងនេះនៅតែមានអតិបរមា។ ក្នុងពេលជាមួយគ្នានេះ ប្រេកង់នៃ alleles បុគ្គលនៅក្នុងចំនួនប្រជាជនផ្លាស់ប្តូរ។ តម្លៃជាមធ្យមនៃចរិតលក្ខណៈប្រែប្រួលនៅក្នុងចំនួនប្រជាជនក៏ផ្លាស់ប្តូរផងដែរ។ នៅក្នុងមួយជំនាន់ជាបន្តបន្ទាប់ ការផ្លាស់ប្តូរបន្តិចម្តងៗរបស់ពួកគេក្នុងទិសដៅជាក់លាក់មួយអាចត្រូវបានតាមដាន។ ទម្រង់នៃការជ្រើសរើសនេះត្រូវបានគេហៅថាការជ្រើសរើសបើកបរ។
ឧទាហរណ៍បុរាណ ការជ្រើសរើសការបើកបរគឺជាការវិវត្តនៃពណ៌នៅក្នុងខែ birch ។ ពណ៌នៃស្លាបរបស់មេអំបៅនេះ ធ្វើត្រាប់តាមពណ៌នៃសំបកឈើដែលគ្របដណ្តប់ដោយ lichen ដែលវាចំណាយពេលពេញមួយថ្ងៃ។ ជាក់ស្តែង ពណ៌ការពារបែបនេះត្រូវបានបង្កើតឡើងក្នុងជំនាន់ជាច្រើននៃការវិវត្តន៍ពីមុន។ ទោះជាយ៉ាងណាក៏ដោយជាមួយនឹងការចាប់ផ្តើមនៃបដិវត្តន៍ឧស្សាហកម្មនៅក្នុងប្រទេសអង់គ្លេសឧបករណ៍នេះបានចាប់ផ្តើមបាត់បង់សារៈសំខាន់របស់វា។ ការបំពុលបរិយាកាសបាននាំឱ្យមានការស្លាប់ដ៏ធំនៃ lichens និងការងងឹតនៃគល់ឈើ។ មេអំបៅស្រាលប្រឆាំងនឹងផ្ទៃខាងក្រោយងងឹត បានក្លាយជាសត្វស្លាបដែលអាចមើលឃើញយ៉ាងងាយស្រួល។ ចាប់ផ្តើមនៅពាក់កណ្តាលសតវត្សទី 19 ទម្រង់នៃមេអំបៅងងឹតដែលផ្លាស់ប្តូរ (melanistic) បានចាប់ផ្តើមលេចឡើងនៅក្នុងចំនួនប្រជាជនខែ birch ។ ប្រេកង់របស់ពួកគេបានកើនឡើងយ៉ាងឆាប់រហ័ស។ នៅចុងបញ្ចប់នៃសតវត្សទី 19 ប្រជាជននៅទីក្រុងមួយចំនួននៃសត្វកណ្ដៀរមានទម្រង់ងងឹតស្ទើរតែទាំងស្រុង ខណៈដែលប្រជាជននៅតាមជនបទបន្តត្រូវបានគ្របដណ្ដប់ដោយទម្រង់ពន្លឺ។ បាតុភូតនេះត្រូវបានគេហៅថា melanism ឧស្សាហកម្ម . អ្នកវិទ្យាសាស្ត្របានរកឃើញថា នៅតំបន់ដែលមានការបំពុល សត្វស្លាបទំនងជាបរិភោគទម្រង់ពណ៌ស្រាល ហើយនៅតំបន់ស្អាត សត្វងងឹត។ ការណែនាំនៃការរឹតបន្តឹងការបំពុលបរិយាកាសនៅក្នុងទសវត្សរ៍ឆ្នាំ 1950 បណ្តាលឱ្យការជ្រើសរើសធម្មជាតិផ្លាស់ប្តូរទិសដៅម្តងទៀត ហើយភាពញឹកញាប់នៃទម្រង់ងងឹតនៅក្នុងប្រជាជនទីក្រុងបានចាប់ផ្តើមធ្លាក់ចុះ។ ពួកគេស្ទើរតែកម្រដូចសព្វថ្ងៃនេះ ដូចកាលពីមុនបដិវត្តន៍ឧស្សាហកម្ម។
ការជ្រើសរើសការបើកបរនាំមកនូវសមាសភាពហ្សែននៃចំនួនប្រជាជនស្របនឹងការផ្លាស់ប្តូរបរិយាកាសខាងក្រៅ ដើម្បីឱ្យកាយសម្បទាមធ្យមរបស់ប្រជាជនត្រូវបានពង្រីកជាអតិបរមា។ នៅលើកោះ Trinidad ត្រី guppy រស់នៅក្នុងទឹកផ្សេងៗគ្នា។ ភាគច្រើននៃអ្នករស់នៅតាមដងទន្លេ និងស្រះទឹក បានស្លាប់ដោយសារធ្មេញរបស់ត្រីមច្ឆា។ នៅតំបន់ខ្ពង់រាប ជីវិតរបស់សត្វក្រួចគឺមានភាពស្ងប់ស្ងាត់ជាង - មានសត្វមំសាសីតិចតួចនៅទីនោះ។ ភាពខុសគ្នាទាំងនេះនៅក្នុងលក្ខខណ្ឌខាងក្រៅនាំឱ្យការពិតដែលថា guppies "កំពូល" និង "បាត" បានវិវត្តទៅជា ទិសដៅផ្សេងគ្នា. "សត្វទាប" ក្រោមការគំរាមកំហែងឥតឈប់ឈរនៃការសម្លាប់រង្គាលចាប់ផ្តើមបន្តពូជនៅអាយុមុននិងបង្កើតកូនតូចៗជាច្រើន។ ឱកាសនៃការរស់រានមានជីវិតសម្រាប់ពួកវានីមួយៗមានតិចតួចណាស់ ប៉ុន្តែមានច្រើន ហើយពួកគេខ្លះអាចបន្តពូជបាន។ "ភ្នំ" ឈានដល់ភាពចាស់ទុំខាងផ្លូវភេទនៅពេលក្រោយ ភាពមានកូនរបស់ពួកគេទាបជាង ប៉ុន្តែកូនចៅរបស់ពួកគេធំជាង។ នៅពេលដែលអ្នកស្រាវជ្រាវបានផ្ទេរ guppies "ការលូតលាស់ទាប" ទៅអាងស្តុកទឹកដែលគ្មានមនុស្សរស់នៅនៅផ្នែកខាងលើនៃទន្លេ ពួកគេបានសង្កេតឃើញការផ្លាស់ប្តូរបន្តិចម្តងៗនៃប្រភេទនៃការអភិវឌ្ឍន៍ត្រី។ ដប់មួយឆ្នាំបន្ទាប់ពីការផ្លាស់ទី ពួកវាកាន់តែធំជាងមុន ចាប់ផ្តើមបង្កាត់ពូជនៅពេលក្រោយ ហើយបង្កើតបានកូនតិចជាងមុន ប៉ុន្តែធំជាង។
អត្រានៃការផ្លាស់ប្តូរប្រេកង់ allele នៅក្នុងចំនួនប្រជាជនមួយ និងតម្លៃមធ្យមនៃលក្ខណៈដែលស្ថិតនៅក្រោមឥទ្ធិពលនៃការជ្រើសរើសគឺអាស្រ័យមិនត្រឹមតែទៅលើអាំងតង់ស៊ីតេនៃការជ្រើសរើសប៉ុណ្ណោះទេ ប៉ុន្តែក៏អាស្រ័យលើរចនាសម្ព័ន្ធហ្សែននៃលក្ខណៈដែលការវិលត្រលប់កើតឡើងផងដែរ។ ការជ្រើសរើសប្រឆាំងនឹងការប្រែប្រួលដែលមានការប្រែប្រួលប្រែទៅជាមានប្រសិទ្ធភាពតិចជាងការប្រឆាំងនឹងអ្នកដែលមានឥទ្ធិពល។ នៅក្នុង heterozygote មួយ allele recessive មិនលេចឡើងនៅក្នុង phenotype ទេហើយដូច្នេះគេចចេញពីការជ្រើសរើស។ ដោយប្រើសមីការ Hardy-Weinberg មនុស្សម្នាក់អាចប៉ាន់ប្រមាណអត្រានៃការផ្លាស់ប្តូរប្រេកង់នៃ allele recessive ក្នុងចំនួនប្រជាជនអាស្រ័យលើអាំងតង់ស៊ីតេនៃការជ្រើសរើស និងសមាមាត្រប្រេកង់ដំបូង។ ប្រេកង់ allele កាន់តែទាប ការលុបបំបាត់របស់វាកាន់តែយឺត។ ដើម្បីកាត់បន្ថយភាពញឹកញាប់នៃការស្លាប់ឡើងវិញពី 0.1 ទៅ 0.05 មានតែ 10 ជំនាន់ប៉ុណ្ណោះដែលត្រូវការ។ 100 ជំនាន់ - ដើម្បីកាត់បន្ថយវាពី 0.01 ទៅ 0.005 និង 1000 ជំនាន់ - ពី 0.001 ទៅ 0.0005 ។
ទម្រង់នៃការជ្រើសរើសធម្មជាតិដើរតួនាទីយ៉ាងសំខាន់ក្នុងការសម្របខ្លួននៃសារពាង្គកាយរស់នៅទៅនឹងលក្ខខណ្ឌខាងក្រៅដែលផ្លាស់ប្តូរតាមពេលវេលា។ វាក៏ធានាផងដែរនូវការចែកចាយយ៉ាងទូលំទូលាយនៃជីវិត ការជ្រៀតចូលរបស់វាទៅក្នុងទីផ្សារអេកូឡូស៊ីដែលអាចធ្វើបានទាំងអស់។ ទោះជាយ៉ាងណាក៏ដោយ វាជាកំហុសក្នុងការគិតដែលថានៅក្នុងលក្ខខណ្ឌស្ថេរភាពនៃអត្ថិភាពនៃការជ្រើសរើសធម្មជាតិឈប់។ នៅក្រោមលក្ខខណ្ឌបែបនេះវាបន្តធ្វើសកម្មភាពក្នុងទម្រង់នៃការជ្រើសរើសស្ថេរភាព។
ការជ្រើសរើសស្ថេរភាព។ការជ្រើសរើសស្ថេរភាពរក្សាស្ថានភាពនៃចំនួនប្រជាជនដែលធានានូវកាយសម្បទាអតិបរមារបស់វាក្រោមលក្ខខណ្ឌអត្ថិភាពថេរ។ ក្នុងជំនាន់នីមួយៗ បុគ្គលដែលងាកចេញពីតម្លៃដ៏ល្អប្រសើរជាមធ្យមសម្រាប់លក្ខណៈប្រែប្រួលត្រូវបានដកចេញ។
ឧទាហរណ៍ជាច្រើននៃសកម្មភាពនៃការជ្រើសរើសស្ថេរភាពនៅក្នុងធម្មជាតិត្រូវបានពិពណ៌នា។ ជាឧទាហរណ៍ នៅ glance ដំបូង វាហាក់បីដូចជាការរួមចំណែកដ៏ធំបំផុតចំពោះហ្សែននៃមនុស្សជំនាន់ក្រោយគួរតែត្រូវបានធ្វើឡើងដោយបុគ្គលដែលមានលទ្ធភាពមានកូនអតិបរមា។ ទោះជាយ៉ាងណាក៏ដោយ ការសង្កេតលើចំនួនប្រជាជនធម្មជាតិនៃសត្វស្លាប និងថនិកសត្វបង្ហាញថា នេះមិនមែនជាករណីនោះទេ។ កូនមាន់ ឬកូននៅក្នុងសំបុកកាន់តែច្រើន វាកាន់តែពិបាកចិញ្ចឹមវា ពួកវានីមួយៗកាន់តែតូច និងខ្សោយ។ ជាលទ្ធផល បុគ្គលដែលមានកូនជាមធ្យមគឺសមបំផុត។
ការជ្រើសរើសឆ្ពោះទៅរកមធ្យមត្រូវបានរកឃើញសម្រាប់លក្ខណៈផ្សេងៗគ្នា។ នៅក្នុងថនិកសត្វ ទារកទើបនឹងកើតទម្ងន់ទាប និងទម្ងន់ខ្ពស់ទំនងជាស្លាប់នៅពេលកើត ឬក្នុងសប្តាហ៍ដំបូងនៃជីវិតជាងទារកទើបនឹងកើតទម្ងន់មធ្យម។ ការសិក្សាអំពីទំហំនៃស្លាបរបស់សត្វស្លាបដែលបានស្លាប់បន្ទាប់ពីព្យុះ បានបង្ហាញថា ភាគច្រើននៃពួកវាមានស្លាបដែលតូចពេក ឬធំពេក។ ហើយនៅក្នុងករណីនេះ បុគ្គលមធ្យមបានប្រែក្លាយជាមនុស្សសម្របខ្លួនខ្លាំងបំផុត។
តើអ្វីជាហេតុផលសម្រាប់រូបរាងថេរនៃទម្រង់ប្រែប្រួលមិនល្អនៅក្នុងលក្ខខណ្ឌថេរនៃអត្ថិភាព? ហេតុអ្វីបានជាការជ្រើសរើសធម្មជាតិមិនអាចលុបបំបាត់ទម្រង់ដែលមិនចង់បានទាំងអស់ម្តងហើយម្តងទៀត? ហេតុផលគឺមិនត្រឹមតែប៉ុណ្ណោះ ហើយមិនច្រើនទេ ការលេចឡើងឥតឈប់ឈរនៃការផ្លាស់ប្តូរថ្មីកាន់តែច្រើនឡើងៗ។ ហេតុផលគឺថា genotypes heterozygous ជាញឹកញាប់សមបំផុត។ នៅពេលដែលឆ្លងកាត់ ពួកវាបំបែកឥតឈប់ឈរ ហើយកូនចៅរបស់ពួកគេបង្កើតកូនចៅដូចគ្នា ជាមួយនឹងការថយចុះនៃកាយសម្បទា។ បាតុភូតនេះត្រូវបានគេហៅថា ប៉ូលីម័រហ្វីសដែលមានតុល្យភាព។
ការជ្រើសរើសផ្លូវភេទ។ឈ្មោលនៃប្រភេទសត្វជាច្រើនបានបង្ហាញយ៉ាងច្បាស់នូវលក្ខណៈផ្លូវភេទបន្ទាប់បន្សំ ដែលមើលដំបូងហាក់ដូចជាមិនប្រែប្រួល៖ កន្ទុយរបស់សត្វក្ងោក រោមដ៏ភ្លឺរបស់បក្សីនៃឋានសួគ៌ និងសត្វសេក សត្វមាន់ក្រហមឆ្អៅ ពណ៌ដ៏គួរឱ្យទាក់ទាញនៃត្រីត្រូពិច ចម្រៀង បក្សី និងកង្កែប ជាដើម។ លក្ខណៈពិសេសទាំងនេះជាច្រើនធ្វើឱ្យស្មុគស្មាញដល់ជីវិតរបស់អ្នកដឹកជញ្ជូនរបស់ពួកគេ និងធ្វើឱ្យពួកវាងាយសម្គាល់ចំពោះសត្វមំសាសី។ វាហាក់ដូចជាថាលក្ខណៈទាំងនេះមិនផ្តល់គុណសម្បត្តិណាមួយដល់ក្រុមហ៊ុនដឹកជញ្ជូនរបស់ពួកគេក្នុងការតស៊ូដើម្បីអត្ថិភាពនោះទេ ហើយពួកគេនៅតែរីករាលដាលយ៉ាងខ្លាំងនៅក្នុងធម្មជាតិ។ តើការជ្រើសរើសតាមធម្មជាតិបានដើរតួនាទីអ្វីខ្លះក្នុងការកើតមាន និងរីករាលដាល?
វាត្រូវបានគេដឹងថាការរស់រានមានជីវិតនៃសារពាង្គកាយគឺជាកត្តាសំខាន់មួយ ប៉ុន្តែមិនមែនជាធាតុផ្សំតែមួយគត់នៃជម្រើសធម្មជាតិនោះទេ។ សមាសធាតុសំខាន់មួយទៀតគឺភាពទាក់ទាញចំពោះបុគ្គលដែលមានភេទផ្ទុយ។ Charles Darwin បានហៅបាតុភូតនេះថាការជ្រើសរើសផ្លូវភេទ។ ដំបូងគាត់បានលើកឡើងពីទម្រង់នៃការជ្រើសរើសនេះនៅក្នុង On the Origin of Species ហើយបន្ទាប់មកបានវិភាគវាយ៉ាងលម្អិតនៅក្នុង The Descent of Man and Sexual Selection។ គាត់ជឿថា "ទម្រង់នៃការជ្រើសរើសនេះមិនត្រូវបានកំណត់ដោយការតស៊ូដើម្បីអត្ថិភាពនៅក្នុងទំនាក់ទំនងនៃសរីរាង្គជាមួយគ្នាទៅវិញទៅមកឬជាមួយ។ លក្ខខណ្ឌខាងក្រៅប៉ុន្តែដោយការប្រកួតប្រជែងរវាងបុគ្គលភេទដូចគ្នា ជាធម្មតាជាបុរសដើម្បីកាន់កាប់បុគ្គលនៃភេទផ្សេងទៀត»។
ការជ្រើសរើសផ្លូវភេទ គឺជាជម្រើសធម្មជាតិ ដើម្បីជោគជ័យក្នុងការបន្តពូជ។ លក្ខណៈដែលកាត់បន្ថយលទ្ធភាពជោគជ័យរបស់ម្ចាស់ផ្ទះអាចលេចឡើង និងរីករាលដាល ប្រសិនបើគុណសម្បត្តិដែលពួកគេផ្តល់សម្រាប់ភាពជោគជ័យនៃការបន្តពូជគឺធំជាងគុណវិបត្តិរបស់ពួកគេសម្រាប់ការរស់រានមានជីវិត។ បុរសដែលមានអាយុខ្លី ប៉ុន្តែត្រូវបានមនុស្សស្រីចូលចិត្ត ហើយបង្កើតកូនច្រើនមានកាយសម្បទាខ្ពស់ជាងបុរសដែលមានអាយុវែង ប៉ុន្តែបង្កើតកូនបានតិចតួច។ នៅក្នុងប្រភេទសត្វជាច្រើន ភាគច្រើននៃបុរសមិនចូលរួមក្នុងការបន្តពូជទាល់តែសោះ។ ក្នុងជំនាន់នីមួយៗ ការប្រកួតប្រជែងគ្នាយ៉ាងខ្លាំងកើតឡើងរវាងបុរសចំពោះស្ត្រី។ ការប្រកួតនេះអាចធ្វើឡើងដោយផ្ទាល់ ហើយបង្ហាញរាងដោយខ្លួនឯងក្នុងទម្រង់នៃការតស៊ូដើម្បីទឹកដី ឬការប្រកួត។ វាក៏អាចកើតឡើងក្នុងទម្រង់ប្រយោល និងត្រូវបានកំណត់ដោយជម្រើសរបស់ស្ត្រី។ ក្នុងករណីដែលស្ត្រីជ្រើសរើសបុរស ការប្រកួតប្រជែងរបស់បុរសបង្ហាញខ្លួនដោយការបង្ហាញរូបរាងដ៏ស្រស់ស្អាត ឬអាកប្បកិរិយាស្មុគ្រស្មាញ។ ស្ត្រីជ្រើសរើសបុរសដែលពួកគេចូលចិត្តបំផុត។ តាមក្បួនមួយទាំងនេះគឺជាបុរសភ្លឺបំផុត។ ប៉ុន្តែហេតុអ្វីបានជាមនុស្សស្រីចូលចិត្តបុរសភ្លឺ?
សម្បទារបស់ស្ត្រីអាស្រ័យលើថាតើនាងអាចវាយតម្លៃកាយសម្បទាជាសក្តានុពលរបស់ឪពុកអនាគតរបស់កូនៗនាងបានដោយគោលបំណង។ នាងត្រូវជ្រើសរើសបុរសដែលកូនប្រុសអាចសម្របខ្លួនបានខ្ពស់ និងទាក់ទាញចំពោះមនុស្សស្រី។
សម្មតិកម្មសំខាន់ពីរអំពីយន្តការនៃការជ្រើសរើសផ្លូវភេទត្រូវបានស្នើឡើង។
យោងតាមសម្មតិកម្ម "កូនប្រុសគួរឱ្យទាក់ទាញ" តក្កវិជ្ជានៃជម្រើសស្ត្រីគឺខុសគ្នាខ្លះ។ ប្រសិនបើបុរសដែលមានពណ៌ភ្លឺ ទោះជាដោយហេតុផលណាក៏ដោយ មានភាពទាក់ទាញចំពោះមនុស្សស្រី នោះគួរតែជ្រើសរើសឪពុកដែលមានពណ៌ភ្លឺសម្រាប់អនាគតកូនប្រុសរបស់គាត់ ព្រោះកូនប្រុសរបស់គាត់នឹងទទួលមរតកហ្សែនពណ៌ភ្លឺ ហើយនឹងមានភាពទាក់ទាញដល់មនុស្សស្រីនៅជំនាន់ក្រោយ។ ដូច្នេះវាមានភាពវិជ្ជមាន មតិកែលម្អដែលនាំឱ្យការពិតដែលថាពីជំនាន់មួយទៅជំនាន់មួយពន្លឺនៃ plumage របស់បុរសកើនឡើងកាន់តែច្រើន។ ដំណើរការនេះបន្តរីកចម្រើនរហូតដល់វាឈានដល់ដែនកំណត់នៃលទ្ធភាពជោគជ័យ។ ចូរយើងស្រមៃមើលស្ថានភាពមួយដែលស្ត្រីជ្រើសរើសបុរសដែលមានកន្ទុយវែង។ ឈ្មោលកន្ទុយវែងបង្កើតកូនបានច្រើនជាងឈ្មោលដែលមានកន្ទុយខ្លី និងមធ្យម ។ ពីមួយជំនាន់ទៅមួយជំនាន់ ប្រវែងនៃកន្ទុយកើនឡើងដោយសារតែស្ត្រីជ្រើសរើសបុរសដែលមិនមានកន្ទុយជាក់លាក់មួយ ប៉ុន្តែមានទំហំធំជាងទំហំមធ្យម។ នៅទីបំផុតកន្ទុយឈានដល់ប្រវែងមួយដែលវាប៉ះពាល់ដល់ភាពរឹងមាំរបស់បុរសគឺមានតុល្យភាពដោយសារភាពទាក់ទាញរបស់វានៅក្នុងភ្នែករបស់ស្ត្រី។
ក្នុងការពន្យល់ពីសម្មតិកម្មទាំងនេះ យើងបានព្យាយាមស្វែងយល់ពីតក្កវិជ្ជានៃសកម្មភាពរបស់បក្សីញី។ វាហាក់បីដូចជាយើងរំពឹងច្រើនពេកពីពួកគេ ដែលការគណនាស្មុគស្មាញនៃកាយសម្បទាបែបនេះគឺស្ទើរតែមិនអាចធ្វើទៅបានសម្រាប់ពួកគេ។ តាមពិតទៅ មនុស្សស្រីមិនមានហេតុផលច្រើន ឬតិចក្នុងជម្រើសរបស់ពួកគេចំពោះបុរសជាងអាកប្បកិរិយាផ្សេងទៀតរបស់ពួកគេទាំងអស់។ នៅពេលដែលសត្វមានអារម្មណ៍ស្រេក វាមិនសមហេតុផលថាវាគួរផឹកទឹកដើម្បីស្ដារតុល្យភាពទឹក-អំបិលក្នុងរាងកាយនោះទេ ពោលគឺវាទៅរន្ធទឹកព្រោះវាមានអារម្មណ៍ស្រេក។ នៅពេលដែលឃ្មុំកម្មករខាំសត្វមំសាសីដែលកំពុងវាយប្រហារសំបុកនោះ នាងមិនបានគណនាថាតើចំនួនប៉ុន្មានជាមួយនឹងការលះបង់ខ្លួនឯងនេះ នាងបានបង្កើនសម្បទារួមរបស់បងប្អូនស្រីរបស់នាង ពោលគឺនាងធ្វើតាមសភាវគតិ។ តាមរបៀបដូចគ្នាស្ត្រីនៅពេលជ្រើសរើសបុរសភ្លឺធ្វើតាមសភាវគតិរបស់ពួកគេ - ពួកគេចូលចិត្តកន្ទុយភ្លឺ។ មនុស្សទាំងអស់ដែលសភាវគតិបានស្នើឱ្យមានអាកប្បកិរិយាខុសគ្នាពួកគេទាំងអស់មិនចាកចេញពីកូនចៅទេ។ ដូច្នេះហើយ យើងកំពុងពិភាក្សាគ្នា មិនមែនជាតក្កវិជ្ជារបស់ស្ត្រីនោះទេ ប៉ុន្តែតក្កវិជ្ជានៃការតស៊ូដើម្បីអត្ថិភាព និងការជ្រើសរើសធម្មជាតិ ដែលជាដំណើរការពិការភ្នែក និងដោយស្វ័យប្រវត្តិ ដែលធ្វើសកម្មភាពឥតឈប់ឈរពីជំនាន់មួយទៅជំនាន់មួយ បានបង្កើតភាពចម្រុះដ៏អស្ចារ្យនៃរូបរាង ពណ៌ និងសភាវគតិដែល យើងសង្កេតឃើញនៅក្នុងពិភពនៃធម្មជាតិរស់នៅ។
4. តួនាទីនៃការប្រែប្រួលតំណពូជក្នុងការវិវត្តន៍នៃប្រភេទសត្វ និងទម្រង់របស់វា
នៅក្នុងទ្រឹស្ដីវិវត្តន៍របស់ដាវីន តម្រូវការជាមុនសម្រាប់ការវិវត្តន៍គឺការប្រែប្រួលតំណពូជ ហើយកម្លាំងជំរុញនៃការវិវត្តន៍គឺជាការតស៊ូដើម្បីអត្ថិភាព និងការជ្រើសរើសធម្មជាតិ។ នៅពេលបង្កើតទ្រឹស្ដីវិវត្តន៍មួយ លោក Charles Darwin បានងាកទៅរកលទ្ធផលនៃការអនុវត្តការបង្កាត់ពូជម្តងហើយម្តងទៀត។ គាត់បានបង្ហាញថាភាពចម្រុះនៃពូជនិងពូជគឺផ្អែកលើការប្រែប្រួល។ ភាពប្រែប្រួលគឺជាដំណើរការនៃការកើតនៃភាពខុសគ្នានៃពូជពង្សបើប្រៀបធៀបទៅនឹងបុព្វបុរស ដែលកំណត់ពីភាពចម្រុះនៃបុគ្គលនៅក្នុងពូជ ឬពូជ។ ដាវីនជឿជាក់ថាមូលហេតុនៃភាពប្រែប្រួលគឺជាផលប៉ះពាល់នៃកត្តាបរិស្ថានលើសារពាង្គកាយ (ដោយផ្ទាល់ និងដោយប្រយោល) ក៏ដូចជាធម្មជាតិនៃសារពាង្គកាយខ្លួនឯង (ចាប់តាំងពីពួកវានីមួយៗមានប្រតិកម្មជាពិសេសចំពោះឥទ្ធិពលនៃបរិយាកាសខាងក្រៅ) មូលដ្ឋានសម្រាប់ការបង្កើតលក្ខណៈថ្មីនៅក្នុងរចនាសម្ព័ន្ធ និងមុខងារនៃសារពាង្គកាយ និងតំណពូជបង្រួបបង្រួមលក្ខណៈទាំងនេះ ដាវីន ការវិភាគទម្រង់នៃភាពប្រែប្រួលបានកំណត់អត្តសញ្ញាណបីក្នុងចំណោមពួកគេ៖ ច្បាស់លាស់ មិនកំណត់ និងជាប់ទាក់ទងគ្នា។
ភាពប្រែប្រួលជាក់លាក់ ឬជាក្រុម គឺជាភាពប្រែប្រួលដែលកើតឡើងក្រោមឥទិ្ធពលនៃកត្តាបរិស្ថានមួយចំនួន ដែលធ្វើសកម្មភាពស្មើៗគ្នាលើបុគ្គលទាំងអស់នៃពូជ ឬពូជ និងការផ្លាស់ប្តូរក្នុងទិសដៅជាក់លាក់មួយ។ ឧទាហរណ៏នៃភាពប្រែប្រួលបែបនេះរួមមានការកើនឡើងនៃទំងន់រាងកាយចំពោះបុគ្គលសត្វដែលមានការចិញ្ចឹមល្អ ការផ្លាស់ប្តូររោមក្រោមឥទ្ធិពលនៃអាកាសធាតុ។ល។ ភាពប្រែប្រួលជាក់លាក់មួយគឺរីករាលដាល គ្របដណ្ដប់លើជំនាន់ទាំងមូល និងត្រូវបានបង្ហាញនៅក្នុងបុគ្គលនីមួយៗតាមរបៀបស្រដៀងគ្នា។ វាមិនមែនជាតំណពូជទេ ពោលគឺនៅក្នុងកូនចៅនៃក្រុមដែលបានកែប្រែក្រោមលក្ខខណ្ឌផ្សេងទៀត លក្ខណៈដែលទទួលបានដោយឪពុកម្តាយមិនត្រូវបានទទួលមរតកទេ។
ភាពប្រែប្រួលមិនច្បាស់លាស់ ឬបុគ្គល បង្ហាញឱ្យឃើញដោយខ្លួនឯងជាពិសេសនៅក្នុងបុគ្គលនីមួយៗ ពោលគឺឧ។ ឯកវចនៈ, បុគ្គលនៅក្នុងធម្មជាតិ។ វាត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងភាពខុសគ្នានៃបុគ្គលដែលមានពូជដូចគ្នា ឬពូជនៅក្រោមលក្ខខណ្ឌស្រដៀងគ្នា។ ទម្រង់នៃភាពប្រែប្រួលនេះគឺមិនប្រាកដប្រជា ពោលគឺលក្ខណៈនៅក្រោមលក្ខខណ្ឌដូចគ្នាអាចផ្លាស់ប្តូរក្នុងទិសដៅផ្សេងៗគ្នា។ ឧទហរណ៍ រុក្ខជាតិមួយប្រភេទ បង្កើតគំរូផ្កាដែលមានពណ៌ខុសៗគ្នា អាំងតង់ស៊ីតេនៃពណ៌នៃផ្កា។ល។ ហេតុផលសម្រាប់បាតុភូតនេះ ដាវីនមិនស្គាល់ទេ។ ភាពប្រែប្រួលមិនច្បាស់លាស់គឺជាតំណពូជនៅក្នុងធម្មជាតិ ពោលគឺវាត្រូវបានបញ្ជូនដោយស្ថេរភាពទៅកូនចៅ។ នេះគឺជាសារៈសំខាន់របស់វាសម្រាប់ការវិវត្តន៍។
ជាមួយនឹងភាពប្រែប្រួល ឬជាប់ទាក់ទងគ្នា ការប្រែប្រួលនៃសរីរាង្គណាមួយបណ្តាលឱ្យមានការផ្លាស់ប្តូរនៅក្នុងសរីរាង្គផ្សេងទៀត។ ជាឧទាហរណ៍ សត្វឆ្កែដែលមានអាវរងារមិនសូវអភិវឌ្ឍ ជាធម្មតាមានធ្មេញមិនសូវលូតលាស់ សត្វព្រាបដែលមានជើងមានរោមមានខ្សែនៅចន្លោះម្រាមជើង ព្រាបដែលមានចំពុះវែងជាធម្មតាមានជើងវែង ឆ្មាពណ៌សដែលមានភ្នែកពណ៌ខៀវជាធម្មតាថ្លង់។ល។ កត្តានៃភាពប្រែប្រួលទាក់ទង ដាវីន ធ្វើការសន្និដ្ឋានដ៏សំខាន់មួយ៖ មនុស្សម្នាក់ជ្រើសរើសលក្ខណៈរចនាសម្ព័ន្ធណាមួយ ស្ទើរតែ "ប្រហែលជាផ្លាស់ប្តូរផ្នែកផ្សេងទៀតនៃរាងកាយដោយអចេតនា ដោយផ្អែកលើច្បាប់អាថ៌កំបាំងនៃការទាក់ទងគ្នា"។
ដោយបានកំណត់ទម្រង់នៃភាពប្រែប្រួល ដាវីនបានសន្និដ្ឋានថា មានតែការផ្លាស់ប្តូរដែលអាចមានបានគឺមានសារៈសំខាន់សម្រាប់ដំណើរការវិវត្តន៍ ដោយសារមានតែពួកវាប៉ុណ្ណោះដែលអាចប្រមូលផ្តុំពីជំនាន់មួយទៅជំនាន់មួយ។ នេះបើតាមលោក Darwin កត្តាចម្បងនៃការវិវត្តន៍ ទម្រង់វប្បធម៌- នេះគឺជាការប្រែប្រួលតំណពូជ និងការជ្រើសរើសដែលធ្វើឡើងដោយមនុស្ស (ដាវីនហៅថាការជ្រើសរើសបែបសិប្បនិម្មិត)។ ភាពប្រែប្រួលគឺជាតម្រូវការជាមុនចាំបាច់សម្រាប់ការជ្រើសរើសសិប្បនិម្មិត ប៉ុន្តែវាមិនកំណត់ពីការបង្កើតពូជ និងពូជថ្មីនោះទេ។
សេចក្តីសន្និដ្ឋាន
ដូច្នេះ ដាវីន ជាលើកដំបូងក្នុងប្រវត្តិសាស្ត្រជីវវិទ្យា បានបង្កើតទ្រឹស្តីនៃការវិវត្តន៍។ នេះគឺជាសារៈសំខាន់នៃវិធីសាស្រ្តដ៏អស្ចារ្យ ហើយធ្វើឱ្យវាអាចធ្វើទៅបានមិនត្រឹមតែដើម្បីបញ្ជាក់អំពីគំនិតនៃការវិវត្តន៍សរីរាង្គយ៉ាងច្បាស់ និងគួរឱ្យជឿជាក់សម្រាប់សហសម័យប៉ុណ្ណោះទេ ប៉ុន្តែថែមទាំងដើម្បីសាកល្បងសុពលភាពនៃទ្រឹស្តីនៃការវិវត្តន៍ខ្លួនឯងផងដែរ។ នេះជាដំណាក់កាលសម្រេចចិត្តមួយក្នុងចំណោមបដិវត្តន៍គំនិតដ៏អស្ចារ្យបំផុតក្នុងវិទ្យាសាស្ត្រធម្មជាតិ។ អ្វីដែលសំខាន់បំផុតនៅក្នុងបដិវត្តន៍នេះគឺការជំនួសគំនិតទ្រឹស្ដីនៃការវិវត្តន៍ជាគំនិតនៃគោលបំណងបឋមជាមួយនឹងគំរូនៃការជ្រើសរើសធម្មជាតិ។ ទោះបីជាមានការរិះគន់យ៉ាងខ្លាំងក្លាក៏ដោយ ទ្រឹស្ដីរបស់ដាវីនបានទទួលការទទួលស្គាល់យ៉ាងឆាប់រហ័ស ដោយសារតែគំនិតនៃការអភិវឌ្ឍន៍ប្រវត្តិសាស្ត្រនៃធម្មជាតិរស់នៅបានពន្យល់ពីការពិតដែលបានសង្កេតប្រសើរជាងគំនិតនៃភាពមិនប្រែប្រួលនៃប្រភេទសត្វ។ ដើម្បីបញ្ជាក់ទ្រឹស្តីរបស់គាត់ ដាវីន មិនដូចអ្នកកាន់តំណែងមុនរបស់គាត់ទេ បានទាញការពិតជាច្រើនដែលមានសម្រាប់គាត់ពីតំបន់ផ្សេងៗគ្នា។ ការបន្លិចនៃទំនាក់ទំនងជីវសាស្រ្ត និងការបកស្រាយអំពីចំនួនប្រជាជនរបស់ពួកគេគឺជាការច្នៃប្រឌិតដ៏សំខាន់បំផុតនៃគំនិតនៃការវិវត្តន៍របស់ដាវីន ហើយផ្តល់សិទ្ធិក្នុងការសន្និដ្ឋានថាដាវីនបានបង្កើតគំនិតផ្ទាល់ខ្លួនរបស់គាត់អំពីការតស៊ូដើម្បីអត្ថិភាព ដែលខុសពីគំនិតរបស់អ្នកកាន់តំណែងមុនរបស់ដាវីន គោលលទ្ធិនៃការវិវត្តន៍នៃពិភពសរីរាង្គ គឺជាទ្រឹស្ដីដំបូងនៃការអភិវឌ្ឍន៍ដែលត្រូវបានបង្កើតឡើង "សម្ភារៈនិយមប្រវត្តិសាស្រ្តធម្មជាតិនៅក្នុងជម្រៅនៃវិទ្យាសាស្រ្តធម្មជាតិ ដែលជាការអនុវត្តដំបូងនៃគោលការណ៍នៃការអភិវឌ្ឍន៍ទៅកាន់វិស័យឯករាជ្យមួយ។ វិទ្យាសាស្រ្តធម្មជាតិ" នេះគឺជាសារៈសំខាន់វិទ្យាសាស្ត្រទូទៅនៃលទ្ធិដាវីន។
គុណសម្បត្តិរបស់ដាវីនស្ថិតនៅក្នុងការពិតដែលថាគាត់បានរកឃើញ កម្លាំងជំរុញការវិវត្តន៍សរីរាង្គ។ ការអភិវឌ្ឍបន្ថែមទៀតនៃជីវវិទ្យាបានធ្វើឱ្យស៊ីជម្រៅ និងបំពេញបន្ថែមគំនិតរបស់គាត់ ដែលបានបម្រើជាមូលដ្ឋានសម្រាប់លទ្ធិដាវីនសម័យទំនើប។ នៅក្នុងមុខវិជ្ជាជីវសាស្រ្តទាំងអស់ កន្លែងឈានមុខគេឥឡូវនេះកាន់កាប់ វិធីសាស្រ្តប្រវត្តិសាស្ត្រការស្រាវជ្រាវដែលអនុញ្ញាតឱ្យមនុស្សម្នាក់សិក្សាពីផ្លូវជាក់លាក់នៃការវិវត្តនៃសារពាង្គកាយ និងជ្រាបចូលយ៉ាងជ្រៅទៅក្នុងខ្លឹមសារនៃបាតុភូតជីវសាស្ត្រ។ ទ្រឹស្ដីវិវត្តន៍របស់ Charles Darwin បានរកឃើញការអនុវត្តយ៉ាងទូលំទូលាយនៅក្នុងទ្រឹស្ដីសំយោគសម័យទំនើប ដែលកត្តាដឹកនាំតែមួយគត់នៃការវិវត្តន៍នៅតែជាជម្រើសធម្មជាតិ ដែលជាសម្ភារៈសម្រាប់ការផ្លាស់ប្តូរ។ ការវិភាគប្រវត្តិសាស្រ្តនៃទ្រឹស្ដីរបស់ដាវីនជៀសមិនរួចផ្តល់នូវបញ្ហាវិធីសាស្រ្តថ្មីនៃវិទ្យាសាស្ត្រដែលអាចក្លាយជាប្រធានបទនៃការស្រាវជ្រាវពិសេស។ ដំណោះស្រាយចំពោះបញ្ហាទាំងនេះរួមបញ្ចូលការពង្រីកផ្នែកចំណេះដឹង ហើយជាលទ្ធផល វឌ្ឍនភាពវិទ្យាសាស្ត្រក្នុងវិស័យជាច្រើន៖ ទាំងផ្នែកជីវវិទ្យា វេជ្ជសាស្ត្រ និងចិត្តវិទ្យា ដែលទ្រឹស្តីវិវត្តន៍របស់ Charles Darwin មានឥទ្ធិពលមិនតិចជាងវិទ្យាសាស្ត្រធម្មជាតិឡើយ។
បញ្ជីអក្សរសិល្ប៍ដែលបានប្រើ
1. Alekseev V.A. មូលដ្ឋានគ្រឹះនៃលទ្ធិដាវីន (ការណែនាំជាប្រវត្តិសាស្ត្រ និងទ្រឹស្តី)។ - អិម, ១៩៦៤ ។
2. Velisov E.A. ឆាល ដាវីន។ ជីវិត ការងារ និងស្នាដៃរបស់ស្ថាបនិកនៃការបង្រៀនវិវត្តន៍។ - អិម, ១៩៥៩ ។
3. Danilova V.S., Kozhevnikov N.N. គំនិតជាមូលដ្ឋាននៃវិទ្យាសាស្ត្រធម្មជាតិ។ – M.: Aspect Press, 2000. – 256 p.
4. Dvoryansky F.A. ដាវីននិយម។ - អិមៈ MSU, 1964. - 234 ទំ។
5. Lemeza N.A., Kamlyuk L.V., Lisov N.D. ការណែនាំសម្រាប់អ្នកដាក់ពាក្យទៅសាកលវិទ្យាល័យ។ – M.: Rolf, Iris-press, 1998. – 496 p.
6. Mamontov S.G. ជីវវិទ្យា៖ មគ្គុទ្ទេសក៍សម្រាប់អ្នកដាក់ពាក្យទៅសាកលវិទ្យាល័យ។ – អិមៈ វិទ្យាល័យ ឆ្នាំ ១៩៩២ – ២៤៥ ទំ។
7. Ruzavin G.I. គំនិតនៃវិទ្យាសាស្ត្រធម្មជាតិទំនើប៖ វគ្គបង្រៀន។ – អិមៈ គម្រោង ឆ្នាំ ២០០២ – ៣៣៦ ទំ។
8. Sadokhin A.P. គំនិតនៃវិទ្យាសាស្ត្រធម្មជាតិទំនើប។ - អិម, ២០០៥ ។
9. Slopov E.F. គំនិតនៃវិទ្យាសាស្ត្រធម្មជាតិទំនើប។ – M.: Vlados, 1999 – 232 p.
10. Smygina S.I. គំនិតនៃវិទ្យាសាស្ត្រធម្មជាតិទំនើប។ - Rostov លេខ 1997 ។
ការពិភាក្សាអំពីបាតុភូត និងដំណើរការនៃភាពប្រែប្រួល និងតំណពូជបង្ហាញថា កត្តាទាំងនេះមានសារៈសំខាន់ណាស់ក្នុងការវិវត្តន៍។ ទោះយ៉ាងណាក៏ដោយពួកគេមិននាំមុខគេទេ។ ការជ្រើសរើសធម្មជាតិមានសារៈសំខាន់ជាចម្បងក្នុងការវិវត្តន៍។
ភាពប្រែប្រួលនៃតំណពូជនៅក្នុងខ្លួនវាមិនកំណត់ "ជោគវាសនា" នៃក្រុមហ៊ុនដឹកជញ្ជូនរបស់វាទេ។ ជាឧទាហរណ៍ ចូរយើងយោងទៅលើការពិតខាងក្រោម។ កញ្ជ្រោងអាក់ទិក (Alopex) កើតឡើងក្នុងទម្រង់តំណពូជពីរ។ បុគ្គលខ្លះមានសមត្ថភាពតំណពូជក្នុងការទទួលបានរោមពណ៌សនៅរដូវរងា។ កញ្ជ្រោងអាក់ទិកបែបនេះត្រូវបានគេហៅថាពណ៌ស។ កញ្ជ្រោងអាក់ទិកផ្សេងទៀតមិនមានសមត្ថភាពនេះទេ។ ទាំងនេះគឺជាអ្វីដែលគេហៅថាកញ្ជ្រោងខៀវ។
វាត្រូវបានបង្ហាញថាទម្រង់ទី 2 មានភាពលេចធ្លោជាងទីមួយក្នុងគុណភាពនេះ ពោលគឺសមត្ថភាពក្នុងការប្រែពណ៌សក្នុងរដូវរងារប្រែទៅជាទ្រព្យសម្បត្តិធ្លាក់ចុះ ហើយការរក្សាពណ៌ងងឹតក្នុងរដូវរងារគឺជាលក្ខណៈលេចធ្លោមួយ។ ការពិតទាំងនេះមិនកំណត់ពីការវិវត្តនៃកញ្ជ្រោងអាក់ទិកទេ។
នៅក្នុងលក្ខខណ្ឌនៃ tundra ទ្វីបនិងនៅលើកោះដែលតភ្ជាប់ដោយទឹកកកទៅដីគោកកញ្ជ្រោងអាកទិកពណ៌សគ្របដណ្តប់ដែលស្មើនឹង 96-97% នៃចំនួនសរុប។ កញ្ជ្រោងខៀវអាកទិកគឺកម្រណាស់នៅទីនេះ។ ផ្ទុយទៅវិញ កញ្ជ្រោងខៀវ ត្រួតត្រាលើកោះ Commander ។ ការពន្យល់ខាងក្រោមសម្រាប់ទំនាក់ទំនងទាំងនេះត្រូវបានស្នើឡើង (Paramonov, 1929)។ នៅក្នុងតំបន់ Tundra ទ្វីប គម្របព្រិលបន្តគ្របដណ្ដប់ ហើយប្រភពអាហារមានកម្រិតខ្លាំងណាស់។ ដូច្នេះមានការប្រកួតប្រជែងខ្លាំងសម្រាប់អាហារទាំងរវាងកញ្ជ្រោងអាក់ទិក និងរវាងសត្វមំសាសីក្រោយៗទៀត និងសត្វមំសាសីផ្សេងទៀតដែលជ្រៀតចូល tundra (កញ្ជ្រោង ចចក និងនៅព្រំដែនជាមួយព្រៃកោងកាង - wolverine) ។ នៅក្រោមលក្ខខណ្ឌទាំងនេះ ពណ៌ការពារពណ៌សផ្តល់នូវគុណសម្បត្តិច្បាស់លាស់ ដែលកំណត់ភាពលេចធ្លោនៃកញ្ជ្រោងអាកទិកពណ៌សនៅក្នុង tundra ទ្វីប។ ទំនាក់ទំនងគឺខុសគ្នានៅលើកោះ Commander (សមុទ្រ Bering) ដែលកញ្ជ្រោងពណ៌ខៀវត្រួតត្រា។ មិនមានគម្របព្រិលដែលបន្ត និងយូរអង្វែងនៅទីនេះទេ អាហារមានច្រើន ហើយការប្រកួតប្រជែងគ្នាកាន់តែខ្សោយ។ ជាក់ស្តែង ភាពខុសគ្នាទាំងនេះនៅក្នុងលក្ខខណ្ឌបរិស្ថានក៏កំណត់ទំនាក់ទំនងជាលេខរវាងទម្រង់ទាំងពីរនៃកញ្ជ្រោងអាក់ទិក ដោយមិនគិតពីភាពលេចធ្លោ ឬភាពប្រែប្រួលនៃពណ៌របស់វា។ ការវិវត្តនៃកញ្ជ្រោងអាក់ទិកត្រូវបានកំណត់ ដូច្នេះមិនត្រឹមតែដោយកត្តាតំណពូជប៉ុណ្ណោះទេ ប៉ុន្តែក្នុងកម្រិតធំជាងនេះដោយទំនាក់ទំនងរបស់វាទៅនឹងបរិស្ថាន ពោលគឺការតស៊ូដើម្បីអត្ថិភាព និងជាលទ្ធផល ការជ្រើសរើសធម្មជាតិ។ កត្តានេះ ដែលជាកត្តាសំខាន់នៃការវិវត្តន៍យ៉ាងដាច់អហង្ការ ចាំបាច់ត្រូវពិចារណាឱ្យបានលម្អិតបន្ថែមទៀត។
ការតស៊ូដើម្បីអត្ថិភាព
ការជ្រើសរើសធម្មជាតិគឺជាកត្តាស្មុគ្រស្មាញដែលកើតឡើងដោយផ្ទាល់ពីទំនាក់ទំនងរវាងសារពាង្គកាយមួយ និងបរិស្ថានជីវសាស្ត្រ និងជីវសាស្ត្រជុំវិញរបស់វា។ ទម្រង់នៃទំនាក់ទំនងទាំងនេះគឺផ្អែកលើការពិតដែលសារពាង្គកាយបង្កើតប្រព័ន្ធឯករាជ្យ ហើយបរិស្ថានបង្កើតប្រព័ន្ធមួយផ្សេងទៀត។ ប្រព័ន្ធទាំងពីរនេះអភិវឌ្ឍដោយផ្អែកលើលំនាំខុសគ្នាទាំងស្រុង ហើយសារពាង្គកាយនីមួយៗត្រូវដោះស្រាយជាមួយនឹងការប្រែប្រួល និងការផ្លាស់ប្តូរលក្ខខណ្ឌបរិស្ថានជានិច្ច។ អត្រានៃការប្រែប្រួល និងការផ្លាស់ប្តូរទាំងនេះតែងតែខ្ពស់ជាងអត្រានៃការផ្លាស់ប្តូរសារពាង្គកាយ ហើយទិសដៅនៃការផ្លាស់ប្តូរបរិស្ថាន និងការប្រែប្រួលនៃសារពាង្គកាយគឺឯករាជ្យពីគ្នាទៅវិញទៅមក។
ដូច្នេះសារពាង្គកាយណាមួយតែងតែទាក់ទងគ្នាទៅនឹងលក្ខខណ្ឌនៃបរិស្ថានដែលខ្លួនវាជាធាតុផ្សំ។ នេះគឺជាកន្លែងដែលទម្រង់នៃទំនាក់ទំនងរវាងសារពាង្គកាយ និងបរិស្ថានរបស់ពួកគេកើតឡើង ដែលដាវីនហៅថាការតស៊ូដើម្បីអត្ថិភាព។ រាងកាយពិតជាត្រូវប្រយុទ្ធជាមួយកត្តាបរិស្ថានរាងកាយ និងគីមី។ ដូច្នេះ ការតស៊ូដើម្បីអត្ថិភាពគឺដូចដែល Engels បានចង្អុលបង្ហាញថា ជាស្ថានភាពធម្មតា និងជាសញ្ញាដែលជៀសមិនរួចនៃអត្ថិភាពនៃទម្រង់រស់នៅណាមួយ។
ទោះជាយ៉ាងណាក៏ដោយអ្វីដែលបាននិយាយខាងលើមិនកំណត់ពីសារៈសំខាន់នៃការវិវត្តន៍នៃការតស៊ូដើម្បីអត្ថិភាពនោះទេ ហើយពីទំនាក់ទំនងដែលបានពិពណ៌នាមិនធ្វើតាមផលវិបាករបស់វាដែលដាវីនចាប់អារម្មណ៍នោះគឺការជ្រើសរើសធម្មជាតិ។ ប្រសិនបើយើងស្រមៃមើលនរណាម្នាក់ ទម្រង់រស់នៅដែលមានស្រាប់នៅក្នុងលក្ខខណ្ឌដែលបានផ្តល់ឱ្យ និងការតស៊ូដើម្បីអត្ថិភាពជាមួយនឹងកត្តារូបវិទ្យា-គីមីនៃបរិស្ថាន បន្ទាប់មកមិនមានផលវិបាកវិវត្តន៍ណាមួយនឹងកើតឡើងពីទំនាក់ទំនងបែបនេះទេ។ ពួកវាកើតឡើងដោយសារតែការពិតនៅក្នុងបរិយាកាសដែលបានផ្តល់ឱ្យវាតែងតែមានចំនួនជាក់លាក់នៃទម្រង់រស់នៅមិនស្មើគ្នា។
វិសមភាពជីវសាស្រ្ត ដូចដែលបានបញ្ជាក់រួចមកហើយ វាកើតចេញពីភាពប្រែប្រួល និងផលវិបាករបស់វា - តំណពូជពូជពង្ស ហេតុអ្វីបានជាបុគ្គលផ្សេងៗគ្នាត្រូវគ្នាទៅនឹងបរិស្ថានដល់កម្រិតផ្សេងៗគ្នា។ ដូច្នេះហើយជោគជ័យរបស់ពួកគេម្នាក់ៗក្នុងការតស៊ូក្នុងជីវិតគឺខុសគ្នា។ នេះគឺជាកន្លែងដែលការស្លាប់ដែលមិនអាចជៀសបាននៃ "សមតិច" និងការរស់រានមានជីវិតនៃ "សមកាន់តែច្រើន" កើតឡើងហើយដូច្នេះការវិវត្តន៍នៃទម្រង់រស់នៅមានភាពប្រសើរឡើង។
ដូច្នេះហើយ សារៈសំខាន់ចម្បងមិនមែនជាទំនាក់ទំនងនៃទម្រង់ការរស់នៅរបស់បុគ្គលម្នាក់ៗជាមួយបរិស្ថាននោះទេ ប៉ុន្តែជាជោគជ័យ ឬបរាជ័យក្នុងការតស៊ូនៃជីវិត ប្រៀបធៀបនឹងជោគជ័យ ឬបរាជ័យក្នុងខ្លួនរបស់បុគ្គលផ្សេងទៀត ដែលតែងតែមានជីវសាស្ត្រមិនស្មើគ្នា ពោលគឺមាន។ ឱកាសនៃការរស់រានមានជីវិតខុសៗគ្នា។ តាមធម្មជាតិ ការប្រកួតប្រជែងកើតឡើងរវាងបុគ្គលម្នាក់ៗ ដែលជាប្រភេទនៃ "ការប្រកួតប្រជែង" ក្នុងការតស៊ូដើម្បីជីវិត។
ទម្រង់មូលដ្ឋាននៃការតស៊ូដើម្បីអត្ថិភាព
ការប្រកួតប្រជែងមានពីរទម្រង់សំខាន់ៗ។
យើងត្រូវបែងចែករវាងការប្រកួតប្រជែងដោយប្រយោល នៅពេលដែលបុគ្គលមិនឈ្លោះគ្នាដោយផ្ទាល់ ប៉ុន្តែប្រើមធ្យោបាយនៃការចិញ្ចឹមជីវិតដូចគ្នាជាមួយនឹងកម្រិតនៃភាពជោគជ័យខុសៗគ្នា ឬទប់ទល់នឹងលក្ខខណ្ឌមិនអំណោយផល និងការប្រកួតប្រជែងដោយផ្ទាល់ នៅពេលដែលទម្រង់ពីរប៉ះទង្គិចគ្នាយ៉ាងសកម្ម។
សម្រាប់ការបំភ្លឺ ដោយប្រយោល។ចូរយើងប្រើឧទាហរណ៍ខាងក្រោម។ Beketova (1896) ។ ក្នុងចំណោមសត្វទន្សាយទាំងពីរ សរសេរថា Beketov ដែលដេញតាមឆ្កែប្រផេះ សត្វដែលលឿនជាង និងរត់គេចពីសត្វទន្សាយនឹងឈ្នះ ប៉ុន្តែតាមទស្សនៈរបស់ Darwinists ទន្សាយដែលរត់ចេញពីការដេញតាម បានប្រយុទ្ធគ្នានៅក្នុង យល់ថាពួកវាប្រែជាមិនស្មើគ្នាខាងជីវសាស្រ្តទាក់ទងនឹងកត្តាបរិស្ថានមួយផ្សេងទៀត - សត្វមំសាសីដែលដេញតាម។ អាស្រ័យហេតុនេះ មានការប្រកួតប្រជែងដោយប្រយោលរវាងពួកគេ។ ក្រោយមកទៀតគឺជាទម្រង់ទូទៅនៃការតស៊ូដើម្បីអត្ថិភាព។
សូមលើកឧទាហរណ៍មួយទៀត។ Bison បានរស់នៅយូរមកហើយនៅ Belovezhskaya Pushcha ។ ក្រោយមកសត្វក្តាន់ក្រហមត្រូវបានគេនាំចូលទៅក្នុងព្រៃនៃ Pushcha ដែលបង្កាត់ពូជនៅទីនេះ បរិមាណដ៏ច្រើន។. សត្វក្តាន់អាចស៊ីស្លឹក និងសំបកដើមឈើតូចៗបាន។ ជាលទ្ធផល ពួកវាបានបំផ្លាញការលូតលាស់របស់ក្មេងដែលជ្រុះចេញយ៉ាងច្រើន ហើយការលូតលាស់វ័យក្មេងរបស់ coniferous បានលេចឡើងនៅកន្លែងដដែលដែលវាធ្លាប់មានពីមុនមក។ ដូច្នេះទិដ្ឋភាពទូទៅនៃ Pushcha បានផ្លាស់ប្តូរ។ នៅកន្លែងដែលព្រៃលិចទឹកធ្លាប់ដុះលូតលាស់ មានសំណើមច្រើន អូរ និងប្រភពទឹក; ជាមួយនឹងការបំផ្លិចបំផ្លាញនៃក្រាស់ deciduous បរិមាណសំណើមស្ទ្រីមនិងប្រភពទឹកបានថយចុះ។ ការផ្លាស់ប្តូរទេសភាពបានជះឥទ្ធិពលដល់ស្ថានភាពទូទៅនៃហ្វូង bison ។ ទីមួយ ប៊ីសុនត្រូវបានគេបង្អត់អាហារដើមឈើ ដែលវាងាយស្រួលបរិភោគ។ ទីពីរ ការបំផ្លិចបំផ្លាញនៃព្រៃដែលជ្រុះបាន ដកហូតបាននូវជម្រកដ៏ងាយស្រួលកំឡុងពេលមានកូន និងក្នុងផ្នែកក្តៅបំផុតនៃថ្ងៃ។ ទីបី ការសម្ងួតអាងស្តុកទឹកបានកាត់បន្ថយចំនួនកន្លែងស្រោចទឹក។ ដូច្នេះការប្រមូលផ្តុំនៃ bison នៅសាកសពទឹកមួយចំនួនក្នុងអំឡុងពេលស្រោចទឹកបាននាំឱ្យមាន រីករាលដាលជំងឺ fascioliasis (fasciola hepatica - ជំងឺថ្លើម) និងការស្លាប់ញឹកញាប់នៃសត្វជាពិសេសសត្វវ័យក្មេង។ ជាលទ្ធផលនៃទំនាក់ទំនងដែលបានពិពណ៌នាចំនួនហ្វូងគោបានចាប់ផ្តើមថយចុះ (Kulagin, 1919) ។ ប៊ីសុនត្រូវបាន«ចាញ់ក្នុងការតស៊ូដើម្បីអត្ថិភាព»។ វាច្បាស់ណាស់ថាទម្រង់នៃការប្រកួតប្រជែងរវាងសត្វក្តាន់ និងប៊ីសុន គឺដោយប្រយោល។
ទំនាក់ទំនងខុសគ្នាបន្តិចបន្តួចត្រូវបានគេសង្កេតឃើញនៅក្នុងករណី ត្រង់ជាឧទាហរណ៍ ការប្រកួតប្រជែង នៅពេលដែលប្រភេទសត្វមួយផ្លាស់ប្តូរទីតាំងមួយផ្សេងទៀតយ៉ាងសកម្ម។ ជាឧទាហរណ៍ យោងទៅតាម Formozov (Paramonov, 1929) នៅលើឧបទ្វីប Kola កញ្ជ្រោងកំពុងជំនួសកញ្ជ្រោងអាកទិកគ្រប់ទីកន្លែង។ នៅប្រទេសអូស្ត្រាលី សត្វទន្សោងព្រៃកំពុងផ្លាស់ទីលំនៅសត្វកកេរដែលស៊ីសាច់។ នៅប្រទេសកាណាដា សត្វកន្លាតបានលុកលុយតំបន់នោះ ហើយកំពុងផ្លាស់ទីលំនៅកញ្ជ្រោង។ Dergunov (1928) បានសង្កេតឃើញការប្រកួតប្រជែងដ៏ខ្លាំងក្លាកំឡុងពេលដាក់សំបុកសម្រាប់បែហោងធ្មែញរវាង kestrels, scorches និង jackdaws ជាមួយនឹង kestrel ផ្លាស់ប្តូរពួកវាទាំងពីរ។ នៅតំបន់ steppe នៃទ្វីបអឺរ៉ុប និងអាស៊ី សត្វក្ងាន saker falcon ជំនួស falcon peregrine ទោះបីជាមានសំបុកសមរម្យសម្រាប់ក្រោយក៏ដោយ។ ទំនាក់ទំនងស្រដៀងគ្នានេះត្រូវបានគេសង្កេតឃើញរវាងរុក្ខជាតិ។ អ្នកនិពន្ធនៃបន្ទាត់ទាំងនេះរួមជាមួយ S.N. Yaguzhinsky បានធ្វើការពិសោធន៍ដូចខាងក្រោម (នៅស្ថានីយ៍ជីវសាស្រ្ត Bolshevskaya ជិតទីក្រុងម៉ូស្គូ) ។ តំបន់នោះដុះពេញដោយស្មៅព្រៃ ត្រូវបានគេកាប់ឆ្ការ និងសាបព្រោះដោយគ្រាប់រុក្ខជាតិដាំដុះ។ ចម្ងាយប្រហែល 30 ម៉ែត្រពីតំបន់នេះ មានដីមួយកន្លែងដាំដោយផ្កាកូលាប។ នៅឆ្នាំបន្ទាប់មិនមានរុក្ខជាតិដាំដុះតែមួយនៅលើទីតាំងសាកល្បងទេ។ ទោះជាយ៉ាងណាក៏ដោយ គម្របស្មៅមិនដំណើរការឡើងវិញទេ បើទោះបីជាគេហទំព័រខ្លួនឯងត្រូវបានកាត់ចេញពីវាក៏ដោយ។ វាបានប្រែក្លាយទាំងអស់ដោយ clover ទោះបីជា clover លូតលាស់នៅចម្ងាយ 30 ម៉ែត្រពីវាក៏ដោយ។ ជាការពិតណាស់គ្រាប់ពូជទាំង clover និងធញ្ញជាតិបានធ្លាក់ចុះនៅលើគេហទំព័រប៉ុន្តែ clover បានជំនួសធញ្ញជាតិ។ ការ៉េមុតស្រួចនៃ clover ឈរប្រឆាំងនឹងផ្ទៃខាងក្រោយធញ្ញជាតិពណ៌បៃតង។
ប្រសិនបើយើងអាចបែងចែករវាងទម្រង់នៃការប្រកួតប្រជែងទាំងពីរនេះ នោះយើងគួរចងចាំថានៅក្នុងស្ថានភាពធម្មជាតិ ការប្រកួតប្រជែងដោយផ្ទាល់ និងដោយប្រយោលត្រូវបានទាក់ទងគ្នា ហើយការបំបែករបស់ពួកគេគឺមានលក្ខខណ្ឌ។
សូម្បីតែនៅក្នុងឧទាហរណ៍បុរាណនៃការប្រកួតប្រជែងជីវិតដោយផ្ទាល់ក៏ដោយ ធាតុផ្សំនៃការប្រកួតប្រជែងដោយប្រយោលតែងតែត្រូវបានបញ្ចូលទៅក្នុងវា ដោយបង្ហាញក្នុងកម្រិតផ្សេងៗគ្នានៃភាពប្រែប្រួលនៃទម្រង់ប្រកួតប្រជែងទៅនឹងលក្ខខណ្ឌបរិស្ថាន។ ជាឧទាហរណ៍ដើម្បីបញ្ជាក់ពីអ្វីដែលបាននិយាយ សូមពិចារណាពីទំនាក់ទំនងរវាងសត្វកណ្តុរពីរប្រភេទ - សត្វកណ្ដុរ (Rattus norvegicus) និងកណ្តុរខ្មៅ ( Rattus rattus) នៅដើមសតវត្សទី 18 កណ្តុរខ្មៅបានគ្រប់គ្រងទ្វីបអឺរ៉ុប។ ទោះជាយ៉ាងណាក៏ដោយ ជាក់ស្តែងប្រហែលឆ្នាំ 1827 ប៉ាស៊ីយូកបានចូលទៅក្នុងទ្វីបអឺរ៉ុប ហើយបានរីករាលដាលយ៉ាងឆាប់រហ័សនៅក្នុងប្រទេសរុស្ស៊ីនៅអឺរ៉ុប។ នៅជុំវិញឆ្នាំ 1730 pasyuk ត្រូវបាននាំយកនៅលើកប៉ាល់ពី East Indies ទៅកាន់ប្រទេសអង់គ្លេស ហើយពីទីនេះវាបានជ្រាបចូលទៅក្នុងទ្វីបអឺរ៉ុបខាងលិច។ ទំនាក់ទំនងរវាងប្រភេទសត្វទាំងនេះជាធម្មតាត្រូវបានកំណត់ដោយការប្រកួតប្រជែងដោយផ្ទាល់។ Pasyuk យ៉ាងសកម្មផ្លាស់ទីកណ្តុរខ្មៅដោយវាយប្រហារវា។ ឧត្តមភាពរបស់វាយោងទៅតាម Brauner (1906) ត្រូវបានកំណត់ដោយហេតុផលដូចខាងក្រោម។
1. Pasyuk ធំជាងនិងខ្លាំងជាង។ គាត់មានកម្ពស់ខ្ពស់ និងវែងជាងកណ្តុរខ្មៅបន្តិច។ ជើងរបស់គាត់កាន់តែក្រាស់ ខ្នងរបស់គាត់កាន់តែធំ។ ឆ្អឹង Pasyuk កាន់តែរឹងមាំ ចំណុចភ្ជាប់សាច់ដុំកាន់តែច្បាស់ ដែលបង្ហាញពីការលូតលាស់សាច់ដុំកាន់តែច្រើន។
2. Pasyuk ហែលបានល្អ ហើយនៅលើទឹកបានយូរជាងកណ្តុរខ្មៅ 3-4 ដង។
3. Pasyuki តែងតែជាផ្នែកវាយលុក និងមានភាពឆេវឆាវខ្លាំង ខណៈកណ្តុរខ្មៅបានត្រឹមការពារខ្លួនប៉ុណ្ណោះ។ មានករណីដែលគេស្គាល់ថាប៉ាយូកវាយប្រហារសូម្បីតែមនុស្ស។
4. Pasyuks មានសភាវគតិហ្វូងសត្វដែលមានការអភិវឌ្ឍន៍ខ្ពស់ ហើយនៅក្នុងការប្រយុទ្ធជាមួយកណ្តុរខ្មៅ ពួកគេជួយគ្នាទៅវិញទៅមក ខណៈដែលកណ្តុរខ្មៅតែងតែប្រយុទ្ធតែម្នាក់ឯង។
ដូច្នេះគុណសម្បត្តិមួយចំនួនកំណត់លទ្ធផលនៃការតស៊ូដែលដូចដែលអាចមើលឃើញពីខាងលើមានធម្មជាតិនៃការប្រកួតប្រជែងដោយផ្ទាល់រវាងប្រភេទសត្វទាំងនេះ។ ជាលទ្ធផលតំបន់ចែកចាយរបស់កណ្តុរខ្មៅត្រូវបានកាត់បន្ថយយ៉ាងខ្លាំងហើយបំបែកនៅក្នុងផ្នែកអ៊ឺរ៉ុបនៃសហភាពសូវៀតទៅជាតំបន់ដាច់ស្រយាលចំនួនបួន (Kuznetsov) ។ ការកាត់បន្ថយ និងការបែកខ្ញែកនៃជួរនេះគឺជាភស្តុតាងនៃស្ថានភាពធ្លាក់ទឹកចិត្តនៃប្រភេទសត្វ។
ទោះយ៉ាងណាក៏ដោយ ទំនាក់ទំនងទាំងនេះមិនអាចនិយាយជារួមបានទេ។ ដូច្នេះយោងទៅតាម Brauner (1906) និង Gamaleya (1903) សមាមាត្រខាងក្រោមត្រូវបានគេរកឃើញនៅក្នុងច្រក Odessa: ក្នុងចំណោមសត្វកណ្តុរដុតចំនួន 24,116 សត្វប៉ាស៊ីយូគីមានចំនួន 93,3% ឥណ្ឌា (ប្រភេទរងខ្មៅ) - មានតែ 3 គំរូប៉ុណ្ណោះ។ ទោះជាយ៉ាងណាក៏ដោយនៅលើកប៉ាល់បរទេសនិងជនជាតិស្បែកសដែលបានមកដល់កំពង់ផែ Odessa ទំនាក់ទំនងគឺខុសគ្នា: ក្នុងចំណោម 735 បំណែកអេហ្ស៊ីប (ខ្មៅ) - 76%; ពណ៌ខ្មៅធម្មតា - 15,5%, ក្រហម (ប្រភេទរងនៃខ្មៅ) - 55 បំណែក, pasyukov - មានតែគំរូពីរប៉ុណ្ណោះ។ Pasyuki, Gamaleya ចង្អុលបង្ហាញថាមានតែនៅក្នុងច្រក Odessa ប៉ុណ្ណោះ។ Brauner ចង្អុលបង្ហាញថាជាក់ស្តែងនៅក្នុងប្រទេសអេហ្ស៊ីប pasyuk មិនផ្លាស់ប្តូរកណ្តុរខ្មៅ (ពូជរបស់វាពោលគឺកណ្តុរអេហ្ស៊ីប) យ៉ាងងាយស្រួលដូចនៅអឺរ៉ុប។ ជាឧទាហរណ៍ ពិតប្រាកដណាស់ ប្រភេទសត្វទាំងពីរនេះមានវត្តមាននៅលើឆ្នេរសមុទ្រអាហ្រ្វិកខាងជើង ហើយទិន្នន័យអំពីសត្វកណ្តុរនៅលើនាវាចំហុយ (សូមមើលខាងលើ) បង្ហាញជាវិជ្ជមានថាការប្រកួតប្រជែងរវាងប្រភេទសត្វទាំងពីរនៅក្នុងលក្ខខណ្ឌនៃឆ្នេរសមុទ្រអាហ្វ្រិកមានលទ្ធផលខុសគ្នា។ សូម្បីតែ Troussard (1905) បានរាយការណ៍ថានៅលើឆ្នេរសមុទ្រអាហ្រ្វិកសត្វកណ្ដុរខ្មៅជ្រៀតចូលភាគខាងត្បូងចូលទៅក្នុងតំបន់វាលខ្សាច់ដែលមិនមានឃ្មុំ។ ដូច្នេះប្រសិនបើនៅអឺរ៉ុប pasyuki ត្រួតត្រានោះនៅអាហ្វ្រិកទំនាក់ទំនងគឺខុសគ្នា។
ការពិតទាំងនេះបង្ហាញថាលទ្ធផលនៃការប្រកួតប្រជែងមិនត្រូវបានកំណត់ដោយគុណសម្បត្តិរូបវន្តនៃប្រភេទសត្វមួយលើប្រភេទមួយផ្សេងទៀតទេហើយវាក៏អាស្រ័យលើកត្តាផ្សេងទៀតផងដែរ - ការសម្របខ្លួនទៅនឹងបរិស្ថានក្នុងន័យទូលំទូលាយនៃពាក្យ។ ដូច្នេះ ការប្រកួតប្រជែងដោយប្រយោល និងដោយផ្ទាល់ ជាក្បួនត្រូវបានភ្ជាប់គ្នាជាទាំងមូល ហើយអាចខុសគ្នាទាំងស្រុងតាមលក្ខខណ្ឌ។
នៅទីនេះវាត្រូវតែត្រូវបានសង្កត់ធ្ងន់ថានៅក្នុងការតស៊ូដើម្បីអត្ថិភាព កត្តា "Malthusian" ពោលគឺ ចំនួនប្រជាជនលើស ប្រាកដជាមានសារៈសំខាន់យ៉ាងច្បាស់លាស់។ ទោះបីជាមិនមែនជាកត្តាចម្បងក៏ដោយ ប៉ុន្តែចំនួនប្រជាជនលើសធ្វើឱ្យការតស៊ូដើម្បីអត្ថិភាពកាន់តែខ្លាំង។ អាំងតង់ស៊ីតេរបស់វាកើនឡើងយ៉ាងខ្លាំង។ មុខតំណែងនេះងាយស្រួលក្នុងការបញ្ជាក់ជាមួយនឹងការពិតដូចខាងក្រោម។ ឧទាហរណ៍ ប្រសិនបើប្រភេទសត្វចូលទៅក្នុងទីជម្រកថ្មី ឬត្រូវបាននាំយកមកទីនេះដោយមនុស្ស នោះក្នុងករណីមួយចំនួន វាត្រូវបានគេសង្កេតឃើញថា វាចាប់ផ្តើមបន្តពូជយ៉ាងខ្លាំងក្លា និងកើនឡើងយ៉ាងឆាប់រហ័សនៅក្នុងចំនួន។ ការសង្កេតបង្ហាញថាបាតុភូតទាំងនេះត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងអវត្តមាននៅក្នុងជម្រកថ្មីនៃដៃគូប្រកួតប្រជែងនិងសត្រូវដែលកាត់បន្ថយចំនួននៃប្រភេទសត្វនេះនៅក្នុងជម្រកពីមុនរបស់វា។
ដូចដែលយើងឃើញ ការតស៊ូដោយប្រយោល និងដោយផ្ទាល់សម្រាប់អត្ថិភាពត្រូវបានភ្ជាប់គ្នាទៅជាស្មុគស្មាញទាំងមូល។ ដូច្នេះ ការយល់ខុសអំពីវាថាជាការតស៊ូដោយផ្ទាល់ក្នុងទម្រង់នៃការប្រយុទ្ធគ្នាដោយផ្ទាល់រវាងសារពាង្គកាយគឺនៅឆ្ងាយបំផុតពីអត្ថន័យពិតនៃពាក្យនេះ។ ផ្ទុយទៅវិញ ការតស៊ូដើម្បីអត្ថិភាពត្រូវតែយល់ក្នុងន័យទូលំទូលាយបំផុត ពោលគឺជាទម្រង់នៃទំនាក់ទំនងផ្ទាល់ និងដោយប្រយោលនៃសារពាង្គកាយជាក់លាក់នីមួយៗចំពោះកត្តាបរិស្ថានជីវសាស្ត្រ និង abiotic ដែលកើតឡើងដោយសារតែទំនាក់ទំនងនៃភាពប្រែប្រួលនៃទម្រង់រស់នៅណាមួយចំពោះណាមួយ។ លក្ខខណ្ឌ និងធាតុផ្សំនៃបរិស្ថាន ក៏ដូចជាដោយសារចំនួនប្រជាជនច្រើនលើសលប់ និងការប្រកួតប្រជែង ដែលកំណត់ការលុបបំបាត់ចោលនូវអ្នកដែលមិនប្រែប្រួល និងការរស់រានមានជីវិតរបស់អ្នកប្រែប្រួល។
ទំនាក់ទំនងស្មុគ្រស្មាញក្នុងការតស៊ូដើម្បីអត្ថិភាព
ឧទាហរណ៍ខាងលើបានពិនិត្យមើលទំនាក់ទំនងដោយផ្ទាល់រវាងប្រភេទសត្វពីរ។ តាមពិតទំនាក់ទំនងនេះគឺស្មុគស្មាញជាង។ ប្រភេទសត្វណាមួយរស់នៅក្នុងតំបន់ជាក់លាក់មួយ ដែលមានជាដំបូង លក្ខណៈរូបវន្ត គីមី អាកាសធាតុ និងទេសភាព។ សីតុណ្ហភាពជាមធ្យមនៅក្នុងតំបន់នេះ បរិមាណទឹកភ្លៀង ចំនួនថ្ងៃច្បាស់លាស់ក្នុងមួយឆ្នាំ ធម្មជាតិ និងកម្រិតនៃភាពស្ងប់ស្ងាត់ ខ្យល់បក់បោក។ សមាសធាតុគីមីដី រចនាសម្ព័ន្ធរូបវន្ត ពណ៌ និងរូបរាងរបស់វា។ ផ្ទៃផែនដីភាពធូរស្រាលរបស់វា ការអវត្ដមាន ឬភាពសម្បូរបែបនៃអាងទឹក - កត្តាទាំងអស់នេះ និងកត្តាផ្សេងទៀតដែលយកមកជាមួយគ្នា គឺជាផ្នែកនៃលក្ខណៈនៃប្រភេទជម្រក ឬស្ថានីយ៍។
ឧទាហរណ៍ ស្ថានីយ៍គឺវាលស្មៅអំបិល វាលស្មៅស្លាប វាលខ្សាច់ថ្ម។ វាលខ្សាច់ព្រៃស្រោង, ព្រៃរបោះ, ព្រៃចម្រុះ (តៃហ្កា) ព្រៃស្រោង, ទនដ្រា។ សម្រាប់សារពាង្គកាយក្នុងទឹកតូចៗ ឬសូម្បីតែមីក្រូទស្សន៍ ស្ថានីយនឹងមានឧទាហរណ៍៖ ខ្សាច់សែល កំរាល Elodea ក្រាស់ Zoster បាត detritus បាតភក់ កន្លែងទឹកបើកចំហ ផ្ទៃថ្មក្រោមទឹក។ល។
រួចទៅហើយពីឧទាហរណ៍ទាំងនេះវាច្បាស់ណាស់ថាស្ថានីយ៍ត្រូវបានបង្កើតឡើងក្រោមឥទិ្ធពលនៃកត្តារូបវិទ្យាមិនត្រឹមតែប៉ុណ្ណោះទេប៉ុន្តែសារពាង្គកាយក៏ចូលរួមក្នុងការបង្កើតរបស់វាផងដែរ (ឧទាហរណ៍ស្ថានីយ៍នៃព្រៃ deciduous) ។ ប៉ុន្តែសារពាង្គកាយសត្វក៏ទុកសញ្ញារបស់វានៅលើស្ថានីយ៍មួយ ហើយសកម្មភាពរបស់ពួកគេក៏កំណត់លក្ខណៈរបស់វាផងដែរ។ គ្រប់សារពាង្គកាយទាំងអស់ដែលរស់នៅក្នុងស្ថានីយ៍ដែលបានផ្តល់ឱ្យគឺស្ថិតនៅក្នុងទំនាក់ទំនងស្មុគ្រស្មាញ ហើយត្រូវបានសម្របតាមលក្ខខណ្ឌរបស់វា។
ចំនួនសរុបនៃទម្រង់ជីវិតនៃស្ថានីយ៍ដែលបានផ្តល់ឱ្យ ដែលស្ថិតនៅក្នុងទំនាក់ទំនងដែលពឹងផ្អែក និងអាស្រ័យគ្នាទៅវិញទៅមក បង្កើតបានជាប្រព័ន្ធអេកូឡូស៊ីដែលបានបង្កើតឡើងជាប្រវត្តិសាស្ត្រនៃទម្រង់ជីវិត (ប្រភេទសត្វ) ឬ biocenosis ។
តួលេខនេះបង្ហាញពី "ខ្សែសង្វាក់អាហារ" ដ៏ស្មុគស្មាញដែលភ្ជាប់ទម្រង់ជីវិតនៃ biocenosis វាលស្មៅ។ ព្រួញចេញពីសត្វព្រៃទៅជាសត្វមំសាសី។ ការផ្លាស់ប្តូរចំនួននៃទម្រង់ជីវិតមួយនាំឱ្យមានការផ្លាស់ប្តូរមួយចំនួននៅក្នុង biocenosis ។ ជាឧទាហរណ៍ ប្រសិនបើសត្វចចកបានសម្លាប់សត្វប្រចៀវ ពួកវាចាប់ផ្តើមស៊ីសត្វកណ្តុរ ក្លាយជាគូប្រជែងរបស់សត្វកន្លាត ដែលប្តូរទៅចិញ្ចឹមសត្វកុក។ ការថយចុះនៃចំនួន gophers នាំឱ្យមានការកើនឡើងនៃចំនួនសត្វល្អិត - កត្តាដែលប៉ះពាល់ដល់បន្លែហើយក្នុងពេលតែមួយអំណោយផលសម្រាប់ទម្រង់ insectivorous ។ល។
ពីខាងលើវាច្បាស់ណាស់ថាទម្រង់ជីវិតដោយផ្ទាល់ឬដោយប្រយោលត្រូវបានរងផលប៉ះពាល់ដោយការផ្លាស់ប្តូរនៅក្នុង biocenosis ។ វាងាយស្រួលក្នុងការយល់ថាការបាត់បង់សមាជិកមួយនៃ biocenosis អាចបណ្តាលឱ្យមានការផ្លាស់ប្តូររ៉ាឌីកាល់នៅក្នុងវា។ តាមពិតនេះគឺជាអ្វីដែលកើតឡើង។ biocenosis ផ្លាស់ប្តូរសមាសភាពរបស់វាតាមពេលវេលា និងវិវត្តទៅជា biocenosis ថ្មី។ ការផ្លាស់ប្តូរនេះនៅក្នុងសមាសភាពនៃ biocenosis ត្រូវបានគេហៅថាបន្តបន្ទាប់។ ការបន្តពូជបង្ហាញយ៉ាងល្អឥតខ្ចោះនូវវត្តមាននៃការតស៊ូសម្រាប់អត្ថិភាពនៅក្នុង biocenosis និងឥទ្ធិពលរបស់វាទៅលើសមាសភាពប្រភេទសត្វ។
សូមក្រឡេកមើលឧទាហរណ៍មួយចំនួន។ ការជំរុញបសុសត្វជាប្រព័ន្ធទៅកាន់វាលស្មៅមួយចំនួននាំទៅដល់ការអភិវឌ្ឍន៍កន្លែងសត្តឃាត។ នៅក្នុងវាលស្មៅ ដំណាក់កាលដំបូងរបស់វា គឺការបំផ្លាញចោលនូវសំណល់រុក្ខជាតិដែលងាប់ ដែលកកកុញពីមួយឆ្នាំទៅមួយឆ្នាំ និងការប៉ះពាល់ដី។ បំណះទំពែកបែបនេះត្រូវបានកាន់កាប់ដោយរុក្ខជាតិប្រចាំឆ្នាំនៃធាតុជនបរទេស។ ដោយសារតែការខ្សោះជីវជាតិនៃ permeability ទឹកនៃដីបង្រួមដោយកន្លែងសត្តឃាតការរីកលូតលាស់នៃស្មៅថយចុះ។ នៅដំណាក់កាលទី 2 ស្មៅរោមនិង tyrsa ថយចុះគួរឱ្យកត់សម្គាល់នៅក្នុងចំនួន fescue ត្រូវបានរក្សាទុកជាបណ្តោះអាសន្នហើយ wormwood, chamomile និង bulbous thinleg ក្លាយជាទម្រង់លេចធ្លោ។ ក្រោយមក ស្មៅរោម និង tyrsa បាត់ទាំងស្រុង ការធ្លាក់ចុះនៃតួលេខ ហើយភាពលេចធ្លោបានឆ្លងទៅ wormwood ជាដើម។ ជាទូទៅ ស្មៅស្មៅដ៏ស្វិតស្វាញត្រូវបានជំនួសដោយស្មៅស្ងួតពាក់កណ្តាលវាលខ្សាច់ដែលមានទឹកដម។ ការផ្លាស់ប្តូរនេះអនុគ្រោះដល់សត្វកកេរ steppe ដែលចំនួនរបស់វាកំពុងកើនឡើងនៅក្នុងតំបន់សត្តឃាត។ ម្យ៉ាងវិញទៀត ការសំលាប់ប៉ះពាល់ដល់សត្វពាហនៈ (សត្វល្អិត)។ Geophilic (ស្រឡាញ់ដី) ទម្រង់ធម្មតានៃស្ថានីយ៍វាលខ្សាច់លេចឡើងឧទាហរណ៍ steppe konik ត្រូវបានជំនួសដោយ prusik ។ល។ (Formozov) ។ ដូចដែលយើងអាចមើលឃើញនៅក្រោមឥទ្ធិពលនៃកត្តាមួយ - ការសំលាប់ - ការបន្តពូជពេញលេញបានកើតឡើងហើយសមាសភាពទាំងមូលនៃ biocenosis បានផ្លាស់ប្តូរ។ របបធារាសាស្ត្រថ្មីនៃដីបានធ្វើឱ្យទម្រង់រុក្ខជាតិពីមុនមិនសម្របទៅនឹងលក្ខខណ្ឌថ្មី ហើយកន្លែងរបស់វាត្រូវបានយកដោយទម្រង់ផ្សេងទៀត ដែលនាំឱ្យមានការផ្លាស់ប្តូរមួយចំនួននៅក្នុងពពួកសត្វ។ ទម្រង់ខ្លះផ្លាស់ទីលំនៅផ្សេងទៀត។
លក្ខណៈពិសេសគួរឱ្យកត់សម្គាល់នៃទំនាក់ទំនងទាំងនេះគឺការពិតដែលថា biocenosis ជាក់លាក់មួយនៅពេលដែលវារីកចម្រើនរៀបចំដើម្បីជំនួសវាជាមួយអ្នកដទៃ។ ជាឧទាហរណ៍ ការចោលសំណល់រុក្ខជាតិនៅលើវាលភក់ស្មៅនាំឲ្យមានការកើនឡើងនៃផ្ទៃវាលភក់។ ជំនួសឱ្យអាងមួយ ការសង្គ្រោះប៉ោងមួយត្រូវបានបង្កើតឡើង។ ការហូរចូលនៃទឹកមានការថយចុះ ហើយជំនួសឱ្យស្មៅ (sedge) bog, sphagnum ដែលមានបន្លែខ្ពស់ជាងនេះ តំណាងដោយ marsh scheuchzeria palustris មានការរីកចម្រើន។ ស្មុគស្មាញនេះ (Sphagnum + Scheuchzeria) ត្រូវបានបង្រួមហើយលក្ខខណ្ឌត្រូវបានបង្កើតឡើងដែលអំណោយផលសម្រាប់ការបន្ថែមទម្រង់ទីបីទៅវា - ស្មៅកប្បាស (Eriophorum yaginatum) ។ ក្នុងពេលជាមួយគ្នានោះគម្រប sphagnum ប្រែទៅជាតំណាងដោយប្រភេទផ្សេងៗគ្នា (ជំនួសឱ្យ Sph. មធ្យម - Sph. Inseni) ។ ការកើនឡើងជាបន្តនៃកំរាលព្រំ sphagnum អនុគ្រោះដល់រូបរាងនៃស្រល់។ ដូច្នេះ biocenosis នីមួយៗរៀបចំការស្លាប់ដោយខ្លួនឯង (Sukachev, 1922) ។
បាតុភូតនៃការបន្តពូជបង្ហាញពីបាតុភូតនៃការតស៊ូដើម្បីអត្ថិភាពនៅក្នុង biocenosis មួយ។
ការប្រែប្រួលនៃចំនួនប្រភេទសត្វដែលជាការបង្ហាញនៃការតស៊ូដើម្បីអត្ថិភាព
ការពិតដ៏សំខាន់មួយទៀតដែលបង្ហាញពីការតស៊ូដើម្បីអត្ថិភាពគឺការប្រែប្រួលនៃចំនួនប្រភេទសត្វជុំវិញវដ្តប្រចាំឆ្នាំ។
ការពិតនេះត្រូវបានសិក្សាទាក់ទងទៅនឹងទម្រង់មួយចំនួន - សត្វកកេរដែលបង្កគ្រោះថ្នាក់ សត្វពាហនៈជាដើម។
តួលេខបង្ហាញថាឆ្នាំនៃការធ្លាក់ទឹកចិត្តជាលេខត្រូវបានបន្តដោយឆ្នាំនៃការកើនឡើងជាលេខ ហើយការប្រែប្រួលនៃចំនួនគឺប្រហាក់ប្រហែលនឹងចង្វាក់ធម្មជាតិ។ ចូរយើងពិចារណាអំពីបាតុភូតនៃ "រលកនៃជីវិត" ដែលទាក់ទងយ៉ាងជិតស្និទ្ធទៅនឹងការតស៊ូដើម្បីអត្ថិភាព។
ក. ហេតុផលសម្រាប់ចង្វាក់នៃការប្រែប្រួលចំនួនប្រជាជន. គេបានរកឃើញថាចង្វាក់នៃការប្រែប្រួលលេខគឺខុសគ្នាសម្រាប់ប្រភេទផ្សេងគ្នា។ ឧទាហរណ៍ សម្រាប់សត្វកកេរដូចកណ្ដុរ វាស្មើនឹងជាមធ្យមដប់ឆ្នាំ (Vinogradov, 1934) សម្រាប់កញ្ជ្រោងអាកទិក 2-4 ឆ្នាំសម្រាប់កំប្រុក។ ព្រៃខាងជើងអឺរ៉ាសៀ និងអាមេរិក អាយុ 8-11 ឆ្នាំ ។ល។ ឆ្នាំនៃការធ្លាក់ទឹកចិត្តជាលេខត្រូវបានបន្តដោយឆ្នាំនៃការងើបឡើងវិញ។ ជាក់ស្តែងហេតុផលសម្រាប់ធម្មជាតិនៃចង្វាក់គឺអាស្រ័យមួយផ្នែកទៅលើលក្ខណៈជាក់លាក់នៃប្រភេទជីវសាស្រ្តនីមួយៗ។ ដូច្នេះ S. A. Severtsov (1941) ចង្អុលបង្ហាញថាប្រភេទសត្វនីមួយៗត្រូវបានកំណត់លក្ខណៈដោយអត្រាមរណភាពបុគ្គលជាក់លាក់។ ដោយសារការមានកូននៃប្រភេទនីមួយៗមានលក្ខណៈជាមធ្យមសម្រាប់វា នេះផ្តល់នូវការកើនឡើងដល់ខ្សែកោងកំណើនប្រជាជនជាក់លាក់។ អត្រាកំណើនអ្នកផលិតកាន់តែទាប ការកើនឡើងនៃចំនួនកាន់តែយឺត (Severtsov, 1941) ។ អាស្រ័យហេតុនេះ ការកើនឡើងនៃចំនួន (បន្តពូជ) កើតឡើងទាក់ទងនឹងប្រភេទសត្វនីមួយៗក្នុងកម្រិតជាក់លាក់មួយតាមធម្មជាតិ។ វាមានរយៈពេលមួយរយៈ ក្នុងអំឡុងពេលដែលដង់ស៊ីតេប្រជាជននៃប្រភេទសត្វកើនឡើងជាលំដាប់ ហើយចំនួនអតិបរមានៃដង់ស៊ីតេនេះគឺខុសគ្នាម្តងទៀតសម្រាប់ទម្រង់ផ្សេងៗគ្នា។ ដូច្នេះសម្រាប់សត្វកណ្តុរវាគឺ 5 លាន។ បំណែកក្នុងមួយ sq ។ ម៉ាយ និងសម្រាប់ទន្សាយ ១០០០ ក្នុងមួយម៉ែត្រការ៉េ។ ម៉ាយ (Severtsov, 1941) ។ នៅពេលឈានដល់ដង់ស៊ីតេប្រជាជនកាន់តែខ្ពស់ កត្តាលុបបំបាត់មិនអំណោយផលមួយចំនួនកើតឡើង។ ទន្ទឹមនឹងនេះទម្រង់ផ្សេងៗគ្នាមានបន្សំផ្សេងៗគ្នានៃកត្តាលុបបំបាត់ដែលភាគច្រើនប៉ះពាល់ដល់ពួកគេ។ សម្រាប់សត្វកកេរ សត្វកកេរដែលកើតឡើងជាលទ្ធផលនៃទំនាក់ទំនងជិតស្និទ្ធរវាងបុគ្គលក្នុងអំឡុងពេលបន្តពូជគឺមានសារៈសំខាន់បំផុត។ នៅក្នុង ungulates, epizootics និងការធ្លាក់ទឹកចិត្តអាកាសធាតុគឺមានសារៈសំខាន់ដ៏អស្ចារ្យ។ ទោះជាយ៉ាងណាក៏ដោយ ជាឧទាហរណ៍ ប៊ីសុន រងផលប៉ះពាល់តិចតួចដោយលក្ខខណ្ឌអាកាសធាតុកាន់តែយ៉ាប់យ៉ឺន (ធន់ទ្រាំនឹងពួកវា) ហើយអេពីហ្សូត ផ្ទុយទៅវិញ មានសារសំខាន់ក្នុងការលុបបំបាត់។ ផ្ទុយទៅវិញ ជ្រូកព្រៃមិនទទួលរងពីជំងឺឆ្កួតជ្រូក ជាដើម (Severtsov, 1941)។ អាស្រ័យហេតុនេះ ពីផ្នែកនេះ ភាពជាក់លាក់នៃប្រភេទសត្វអាចមើលឃើញយ៉ាងច្បាស់ថាជាហេតុផលសម្រាប់ចង្វាក់នៃលំយោល។ នេះក៏ត្រូវបានបញ្ជាក់ដោយការពិតដែលថានៅក្នុងទម្រង់ omnivorous (euryphages) ចង្វាក់នៃការប្រែប្រួលនៃលេខគឺមិនសូវច្បាស់ជាងទម្រង់ដែលភ្ជាប់ទៅនឹងអាហារឯកតា (stenophages) ។ ឧទាហរណ៍នៅក្នុងកញ្ជ្រោង omnivorous ភាពប្រែប្រួលនៃលក្ខខណ្ឌនៃការចិញ្ចឹមមិនបណ្តាលឱ្យមានការប្រែប្រួលយ៉ាងខ្លាំងនៃចំនួន (Naumov, 1938) ។ ផ្ទុយទៅវិញ សម្រាប់សត្វកំប្រុក ទិន្នផលគ្រាប់ពូជគឺចាំបាច់ណាស់។ ដើមឈើ coniferous(Formozov, Naumov និង Kiris, 1934) និងភាពប្រែប្រួលនៃចំនួនរបស់វាគឺមានសារៈសំខាន់។
នៅទីបញ្ចប់សូមឱ្យយើងចង្អុលបង្ហាញថាប្រភេទសត្វនីមួយៗមានសក្ដានុពលជីវសាស្រ្តជាក់លាក់មួយ ដោយ Chapman (1928) យល់ពីកម្រិតនៃការតស៊ូដែលកំណត់ដោយតំណពូជនៃប្រភេទសត្វក្នុងការតស៊ូដើម្បីរស់ កំណត់ដោយសក្តានុពលបន្តពូជ និងសក្តានុពលសម្រាប់ការរស់រានមានជីវិតក្នុងការប្រែប្រួលនៃលក្ខខណ្ឌបរិស្ថាន។ .
ដូច្នេះ ពិតណាស់ សម្រាប់ប្រភេទនីមួយៗ មានចង្វាក់ត្រឹមត្រូវនៃការប្រែប្រួលជាលេខ ដែលកំណត់ដោយសក្តានុពលជីវសាស្ត្ររបស់វា។
ទោះជាយ៉ាងណាក៏ដោយ សារៈសំខាន់នៃកត្តានេះមិនគួរត្រូវបានប៉ាន់ស្មានហួសហេតុនោះទេ។ ហេតុផល "ខាងក្នុង" សម្រាប់ចង្វាក់នៃការប្រែប្រួលជាលេខ ដែលបង្ហាញឱ្យឃើញនៅពេលប្រៀបធៀបប្រភេទសត្វផ្សេងៗគ្នា ត្រូវបានគ្របដណ្តប់ដោយហេតុផល "ខាងក្រៅ" ពោលគឺលក្ខខណ្ឌបរិស្ថាននៅក្នុងប្រភេទនីមួយៗនីមួយៗ។ ជាឧទាហរណ៍ ក្នុងចំណោមសត្វកញ្ជ្រោងដែលរស់នៅក្នុងព្រៃ ភាពប្រែប្រួលនៃចំនួនមិនច្រើនទេ ប៉ុន្តែនៅតំបន់វាលស្មៅ និងវាលខ្សាច់ ពួកវាកាន់តែគួរឱ្យកត់សម្គាល់ (Naumov, 1938) ។ សម្រាប់សត្វកំប្រុកចង្វាក់នៃការប្រែប្រួលលេខនៅក្នុងលក្ខខណ្ឌ ព្រៃខាងជើងអឺរ៉ាសៀ និងអាមេរិក ដូចដែលបានបង្ហាញគឺ 8-11 ឆ្នាំនៅក្នុងរយៈទទឹងកណ្តាល 7 ឆ្នាំ និងនៅតំបន់ភាគខាងត្បូងនៃជួររបស់វា - 5 ឆ្នាំ (Naumov, 1938) ។
ទិន្នន័យទាំងនេះបញ្ជាក់ថានៅក្នុង លក្ខខណ្ឌផ្សេងគ្នាការតស៊ូដើម្បីអត្ថិភាពមានអាំងតង់ស៊ីតេខុសៗគ្នា ហើយវាមិនត្រូវបានកំណត់ត្រឹមតែលក្ខណៈ "ខាងក្នុង" នៃប្រភេទសត្វនោះទេ។ ចំពោះសត្វល្អិត ជាទូទៅមិនអាចបង្កើតចង្វាក់ត្រឹមត្រូវនៃការប្រែប្រួលលេខបានទេ ដូចដែលអាចមើលឃើញពីទិន្នន័យខាងក្រោមសម្រាប់ជាយក្រុងម៉ូស្គូ (Kulagin, 1932)។
ទីបំផុត សំណួរក្នុងគ្រប់ករណីទាំងអស់ត្រូវបានគ្របដណ្តប់ដោយទំនាក់ទំនងរវាងប្រភេទសត្វ និងបរិស្ថាន។
ខ. ធាតុផ្សំនៃសក្តានុពលជីវសាស្ត្រនៃប្រភេទសត្វ. ដូចដែលបានបញ្ជាក់រួចមកហើយ សក្តានុពលជីវសាស្ត្រនៃប្រភេទសត្វគឺស្មុគស្មាញទាំងមូល ដែលរួមមានសក្តានុពលបន្តពូជ និងការរស់រានមានជីវិត។ អនុញ្ញាតឱ្យយើងពិចារណាធាតុទាំងនេះនៃសក្តានុពល biotic ដាច់ដោយឡែក។
សក្តានុពលនៃការបន្តពូជជាដំបូងនៃការទាំងអស់គឺអាស្រ័យលើការមានកូននៃប្រភេទសត្វ។ ក្រោយមកទៀតត្រូវបានកំណត់ដោយចំនួនកូនគោនៅក្នុងទុកដាក់សំរាម និងចំនួននៃការទុកដាក់សំរាមក្នុងមួយឆ្នាំ។ កត្តាទាំងនេះនាំឱ្យមានការកើនឡើងយ៉ាងខ្លាំងនៃចំនួនកូនចៅ។ ជាឧទាហរណ៍ អត្រាបន្តពូជរបស់ចាបគឺដូចនេះ ដោយសន្មត់ថាកូនចៅទាំងអស់នៅរស់រានមានជីវិត ចាបមួយគូក្នុងរយៈពេលដប់ឆ្នាំនឹងបង្កើតបានចំនួនប្រជាជនចំនួន 257,716,983,696 នាក់។ កូនរុយផ្លែមួយគូ ដែលបង្កើតបានជាមធ្យម 30 ក្ដាប់នៃ 40 ពងក្នុងមួយឆ្នាំ នឹងគ្របដណ្ដប់លើផែនដីទាំងមូលជាមួយនឹងស្រទាប់ក្រាស់មួយលានម៉ាយក្នុងមួយឆ្នាំ។ នៅក្រោមលក្ខខណ្ឌដូចគ្នានេះ aphid hop បុគ្គលមួយនឹងបង្កើតកូនចៅរបស់បុគ្គល 1022 នៅរដូវក្តៅ។ ដង្កូវនាងហ្គាម៉ាមួយក្បាលអាចបង្កើតបាន 125,000 ដង្កូវតាមទ្រឹស្តី ក្នុងរយៈពេលរដូវក្តៅ។
ទោះជាយ៉ាងណាក៏ដោយ សក្ដានុពលនៃការបន្តពូជរបស់ប្រភេទសត្វមួយមិនត្រឹមតែអាស្រ័យទៅលើការមានកូនប៉ុណ្ណោះទេ។ អាយុនៃការចេញផ្លែដំបូងរបស់ស្ត្រីក៏មានសារៈសំខាន់ខ្លាំងផងដែរ។ ដូចដែល S.A. Severtsov (1941) ចង្អុលបង្ហាញថា ជាមួយនឹងចំនួនកូនគោស្មើគ្នា ជាប្រភេទសត្វដែលញីឈានដល់ភាពពេញវ័យផ្លូវភេទនៅអាយុមុន ហើយដែលរយៈពេលរវាងកំណើតពីរគឺខ្លីជាងនឹងបន្តពូជលឿន។
សារៈសំខាន់ដ៏អស្ចារ្យ បន្ថែមទៀតគឺអាយុសង្ឃឹមរបស់បុគ្គលនៃប្រភេទសត្វ - តម្លៃជាមធ្យមជាក់លាក់ចំពោះប្រភេទនីមួយៗ (S. A. Severtsov, 1941) ។ ដោយមិនគិតពីបញ្ហានេះឱ្យបានលម្អិតទេ យើងនឹងចង្អុលបង្ហាញថាប្រភេទសត្វដែលមានកូនទាបបំផុត ទោះជាយ៉ាងនេះក៏ដោយ អាចមានសក្តានុពលបន្តពូជខ្ពស់ ប្រសិនបើពួកវាត្រូវបានកំណត់លក្ខណៈដោយអាយុជីវិតបុគ្គលដ៏វែង។ ឧទាហរណ៍បុរាណនៃប្រភេទនេះ ជាការយោងរបស់ដាវីនចំពោះការបន្តពូជរបស់ដំរី។ ទោះបីជាមានភាពយឺតយ៉ាវពិសេសនៃការបន្តពូជរបស់វាក៏ដោយ ការគណនាតាមទ្រឹស្តីបង្ហាញថា "ក្នុងកំឡុងឆ្នាំ 740-750 ឆ្នាំ សត្វដំរីមួយគូអាចបង្កើតបានប្រហែលដប់ប្រាំបួនលានដំរីរស់នៅ" (Darwin) ។ ជាចុងក្រោយវាត្រូវតែត្រូវបានសង្កត់ធ្ងន់ថាសក្តានុពលបន្តពូជក៏អាស្រ័យលើលក្ខខណ្ឌនៃការអភិវឌ្ឍនៃកូនចៅនិងជាពិសេសលើទម្រង់នៃការថែទាំសម្រាប់កូនចៅ។ ដោយមិនពឹងផ្អែកលើការពិពណ៌នានៃបាតុភូតខ្លួនវាដែលមានតួអក្សរខុសគ្នាខ្លាំងនៅក្នុងក្រុមផ្សេងៗគ្នានៃសត្វនោះយើងនឹងគ្រាន់តែសង្កត់ធ្ងន់ថាការថែទាំកូនចៅបង្កើនសក្តានុពលនៃការបន្តពូជ។ ដូច្នេះជាក្បួនក្នុងទម្រង់ដែលមានភាពមានកូនទាប មានការអភិវឌ្ឍន៍ខ្លាំងនៃការសម្របខ្លួនដើម្បីការពារកូនចៅ។ ផ្ទុយទៅវិញ អវត្តមាន ឬការបញ្ចេញមតិខ្សោយនៃការសម្របខ្លួនបែបនេះ ជាក្បួនត្រូវបានផ្តល់សំណងដោយការមានកូនខ្ពស់។ ដូច្នេះ សក្ដានុពលនៃការបន្តពូជត្រូវបានកំណត់ដោយកត្តាមួយចំនួន៖ ការមានកូន ចំនួននៃការទុកដាក់ក្នុងមួយឆ្នាំ អាយុសង្ឃឹមរស់ ការសម្របខ្លួនដើម្បីការពារកូនចៅ។
សក្តានុពលរស់រានមានជីវិតគឺជាបរិមាណនៃលំដាប់ផ្សេងគ្នា និងត្រូវបានកំណត់ដោយកម្រិតនៃការសម្របខ្លួនរបស់បុគ្គលនៃប្រភេទសត្វទៅនឹងលក្ខខណ្ឌនៃស្ថានីយ៍របស់ពួកគេ។ សម្បទានេះ ដូចដែលយើងដឹងរួចមកហើយ គឺទាក់ទងគ្នា ដែលជាមូលហេតុដែលកត្តាបរិស្ថានជាច្រើនមានឥទ្ធិពលលើចំនួនប្រជាជននៃប្រភេទសត្វក្នុងលក្ខណៈលុបបំបាត់ (បំផ្លិចបំផ្លាញ) កាត់បន្ថយឥទ្ធិពលនៃសក្តានុពលបន្តពូជ។ តើកត្តាអ្វីខ្លះ ដែលអាចបន្តពូជបានកម្រិតមធ្យម? សូមក្រឡេកមើលពួកគេដោយសង្ខេប។
សារៈសំខាន់ដ៏អស្ចារ្យ ជាដំបូងនៃការទាំងអស់ កត្តាអាកាសធាតុជាពិសេសសីតុណ្ហភាព និងទឹកភ្លៀង។ សម្រាប់ប្រភេទសត្វនីមួយៗ មានកត្តាអាកាសធាតុដ៏ល្អប្រសើរមួយ ដែលអត្រារស់រានមានជីវិតកើនឡើង និងចំនួនប្រភេទសត្វកើនឡើង ស្របតាមសក្តានុពលនៃការបន្តពូជរបស់វា។ តាមធម្មជាតិ ក្នុងរយៈពេលប៉ុន្មានឆ្នាំជិតនឹងលក្ខខណ្ឌដ៏ល្អប្រសើរ ខ្សែកោង "រលកនៃជីវិត" កើនឡើង ហើយផ្ទុយមកវិញ - គម្លាតពីកម្រិតល្អបំផុត ក្នុងទិសដៅមួយ ឬទិសដៅមួយទៀត កាត់បន្ថយការបន្តពូជ។ ចូរយើងផ្តល់ឧទាហរណ៍មួយចំនួន។
ក្នុងរដូវរងារឆ្នាំ 1928 នៅតំបន់ជុំវិញ Leningrad មានការបង្កកដ៏ធំនៃស្ពៃក្តោបពណ៌សនៅក្នុងរដូវរងា ហើយក្នុងរដូវរងាឆ្នាំ 1924/25 - ដង្កូវមេទ័ពធ្លាក់។ វាត្រូវបានបង្កើតឡើងដោយពិសោធន៍ថា ឧទាហរណ៍ ការចិញ្ចឹមកូនឆ្កែរដូវរងានៅ T ° +22.5 ° C បង្កើនការមានកូនរបស់មេអំបៅដែលញាស់ពីពួកវាដល់អតិបរមា (1500-2000 ពង) ។ ទោះជាយ៉ាងណាក៏ដោយ ការប្រែប្រួលក្នុងទិសដៅមួយ ឬទិសដៅមួយទៀតពីកម្រិតល្អបំផុតនេះកាត់បន្ថយការមានកូន។ ដូច្នេះនៅ T ° = +10-12 ° C ភាពមានកូនរបស់មេអំបៅធ្លាក់ចុះដល់ 50% ។ នៅក្នុងសត្វដែលមានឈាមក្តៅ ដោយសារតែសមត្ថភាពគ្រប់គ្រងកំដៅ កត្តាសីតុណ្ហភាពមានឥទ្ធិពលតិចជាង។ ទោះជាយ៉ាងណាក៏ដោយការផ្លាស់ប្តូរសីតុណ្ហភាពនៅតែប៉ះពាល់ដល់ឧទាហរណ៍អត្រានៃការអភិវឌ្ឍន៍នៃ gonads ។ ការកើនឡើងនៃ T° ដល់ដែនកំណត់ជាក់លាក់មួយបង្កើនល្បឿននៃការបង្កើត gonads ទោះជាយ៉ាងណាក៏ដោយការកើនឡើងបន្ថែមទៀតរបស់វាមានឥទ្ធិពលរារាំង។
កត្តាអាកាសធាតុមិនត្រឹមតែប៉ះពាល់ដល់ការមានកូនប៉ុណ្ណោះទេ ប៉ុន្តែក៏មានចំនួនបុគ្គលនៃប្រភេទសត្វផងដែរ។ ជាឧទាហរណ៍ ក្នុងរដូវរងាដ៏អាក្រក់ មានការកើនឡើងនូវភាគរយនៃការស្លាប់របស់សត្វ។ ទិន្នន័យគួរឱ្យចាប់អារម្មណ៍អំពីការស្លាប់របស់សត្វស្លាបក្នុងរដូវរងាធ្ងន់ធ្ងរនៃឆ្នាំ 1939/1940 ត្រូវបានរាយការណ៍ដោយ Dementyev និង Shimbireva (1941) ។ ជាឧទាហរណ៍ បន្ទះប្រផេះមាននៅកន្លែងខ្លះស្ទើរតែស្លាប់ទាំងស្រុង ឬបានថយចុះយ៉ាងខ្លាំងនៅក្នុងចំនួន។ សង្កេត ការស្លាប់ដ៏ធំ coots, waterfowl ជាច្រើន, សត្វទីទុយ (នៅអ៊ុយក្រែន), ចាប, bullfinches, redpolls, siskins, crossbills ជាដើម។
ការលុបបំបាត់ឥទ្ធិពលនៃកត្តាអាកាសធាតុមានលក្ខណៈពីរ (ដោយផ្ទាល់និងដោយប្រយោល) ប៉ះពាល់ដល់ឧទាហរណ៍អាហារូបត្ថម្ភ (បរិមាណចំណី) និងភាពធន់ទ្រាំទៅនឹងជំងឺ (ការចុះខ្សោយនៃរាងកាយ) ។
នៅជាប់នឹងអាកាសធាតុគួរតែត្រូវបានដាក់ដីឬ កត្តា edaphic. នៅក្នុងឆ្នាំស្ងួតដីត្រូវបានបាត់បង់សំណើមច្រើនឬតិចហើយបាតុភូតនេះមានឥទ្ធិពលមធ្យមលើការបន្តពូជរបស់សត្វល្អិតជាច្រើនដំណាក់កាលដង្កូវដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងជីវសាស្រ្តជាមួយដី។ ការត្រជាក់នៃដីក្នុងរដូវរងារក៏បំផ្លាញទម្រង់ជាច្រើនផងដែរ។
Predators មានឥទ្ធិពលសម្របសម្រួលយ៉ាងខ្លាំងលើការបន្តពូជ។ ក្នុងករណីខ្លះវាស្ទើរតែសម្រេចចិត្ត។ ឧទាហរណ៍ សត្វល្អិត Vedalia cardinalis រំខានដល់ការបន្តពូជរបស់សត្វល្អិតខ្នាតធំពី genus Icerya ជាមួយនឹងល្បឿនដ៏អស្ចារ្យដោយសារតែភាពស្អិតរមួតនៃដង្កូវរបស់សត្វល្អិតនេះ និងទម្រង់ពេញវ័យ។ ដង្កូវ vedalia មួយអាចបំផ្លាញដង្កូវមេអំបៅជាង 200 ក្នុងមួយជីវិតរបស់វា។ សត្វល្អិតដីមួយចំនួនក៏ជាភ្នាក់ងារបំផ្លិចបំផ្លាញដ៏មានឥទ្ធិពលផងដែរ។ ការសង្កេតរបស់សត្វកណ្ដៀរដី Carabus nemoralis បានបង្ហាញនូវភាពក្រអឺតក្រទមនៃសត្វកណ្ដៀវដែលចាប់បាននេះ។ ឧទាហរណ៍ ស្ត្រីម្នាក់នៅពេលចាប់បានមានទម្ងន់ ៥៥០ មីលីក្រាម ហើយក្រោយពេលញ៉ាំអាហារ ២,៥ ម៉ោងមានទម្ងន់១០០៥មីលីក្រាម ហើយពោះនាងហើម ហើយលេចចេញពីក្រោមអេលីត្រា។ ការបន្តពូជរបស់សត្វល្អិតក៏ត្រូវបានសម្របសម្រួលដោយបក្សី និងថនិកសត្វផងដែរ។ សត្វស្លាប Insectivorous មានសារសំខាន់ក្នុងរឿងនេះ។ នៅក្នុងព្រៃឈើមួយ វាត្រូវបានគេរកឃើញថា ក្បាលដោះត្រូវបានបំផ្លាញក្នុងរដូវរងាររហូតដល់ 74% នៃដង្កូវនាងមេអំបៅមាសដែលគ្របដណ្តប់លើរដូវរងាទាំងអស់។ ការបំផ្លាញសត្វកកេរដូចសត្វកណ្ដុរដោយសត្វស្លាប និងថនិកសត្វក៏សំខាន់ផងដែរ។ ដូច្នេះ ជាឧទាហរណ៍ ការបំផ្លិចបំផ្លាញនៃ steppe ferret (Putorius eversmanni) បណ្តាលឱ្យមានការកើនឡើងនៃចំនួនសត្វកកេរ។
នៅកន្លែងដែលមានសត្វកកេរប្រមូលផ្តុំ មំសាសីក៏ប្រមូលផ្តុំផងដែរ ដែលរួមចំណែកដល់ការថយចុះចំនួនសត្វកកេរ។ ទំនាក់ទំនងទាំងនេះត្រូវបានកំណត់លក្ខណៈដោយលក្ខណៈពិសេសគួរឱ្យចាប់អារម្មណ៍។ សត្វកកេរដែលរស់នៅក្នុងទីជម្រកបើកចំហច្រើនត្រូវបានសម្លាប់មុន។ នៅក្នុងជម្រកដែលអំណោយផលបំផុតសម្រាប់ការរស់រានមានជីវិត ការស្លាប់របស់សត្វកកេរគឺតិចជាង ហើយពួកវាមិនត្រូវបានបំផ្លាញដោយមំសាសីទេ។ "ស្ថានីយ៍បទពិសោធន៍" បែបនេះ (Naumov, 1939) ដើរតួនាទីនៃទុនបំរុងធម្មជាតិដែលក្នុងនោះសត្វកកេរមិនអាចចូលទៅដល់សត្វមំសាសីបានទេ។ ចំនួនសត្វមំសាសីចាប់ផ្តើមធ្លាក់ចុះ ហើយចំនួនសត្វកកេរចាប់ផ្តើមកើនឡើងស្របតាមសក្តានុពលបន្តពូជជាក់លាក់របស់វា។
ជាទូទៅ ភាពអាស្រ័យនៅទីនេះប្រហាក់ប្រហែលនឹងទំនាក់ទំនងដែលបង្ហាញក្នុងរូប។ ការកើនឡើងនៃចំនួនសត្វឈ្មោលបណ្តាលឱ្យមានការកើនឡើងនៃចំនួនសត្វមំសាសី ហើយការថយចុះជាបន្តបន្ទាប់នៃចំនួនសត្វឈ្មោលកាត់បន្ថយចំនួនសត្វមំសាសី។ សម្រាប់ ប្រភេទបុគ្គលទោះយ៉ាងណាក៏ដោយ ទំនាក់ទំនងលេខស្មុគ្រស្មាញខ្លាំងត្រូវបានគេសង្កេតឃើញ ដែលយើងនឹងពិនិត្យមើលនៅទីនេះក្នុងន័យសង្ខេបបំផុត។
លទ្ធផលនៃសកម្មភាពកម្ចាត់សត្វឈ្មោលគឺអាស្រ័យលើលក្ខណៈរបស់សត្វឈ្មោល លក្ខណៈជាក់លាក់របស់មំសាសី និងលក្ខខណ្ឌបរិស្ថាន។ នៅក្នុងស្ថានភាពលំបាកនៃ biocenosis បញ្ហាត្រូវបានដោះស្រាយជាមួយនឹងការលំបាកយ៉ាងខ្លាំង។ នៅក្នុងការងារមួយចំនួន Gause បានយកផ្លូវនៃការបំបែកបញ្ហា។ ដោយបានជ្រើសរើស ciliates ជាវត្ថុមួយ Gause បានបង្កើតដោយសិប្បនិម្មិតនូវ "មីក្រូកូស" មានកំណត់ ដែលមានឧទាហរណ៍ សត្វពីរប្រភេទ - មំសាសី និងសត្វព្រៃ។ ciliates ពីរត្រូវបានគេយក - Paramaecium caudatum (សត្វព្រៃ) និង Didinium nasutum (អ្នកប្រមាញ់) ។ Didinium ហែលយ៉ាងលឿន (លឿនជាង paramecia) ហើយជញ្ជក់ជនរងគ្រោះរបស់វា។ ដូច្នេះនៅក្នុង "មីក្រូកូស" ដែលមានលក្ខណៈដូចគ្នា ពោលគឺនៅក្នុងឧបករណ៍ផ្ទុកសារធាតុចិញ្ចឹមដោយគ្មាន "ជម្រក" សត្វមំសាសីនៅទីបំផុតបំផ្លាញប៉ារ៉ាមេស៊ីមទាំងស្រុង ហើយស្លាប់ដោយខ្លួនវាផ្ទាល់។ លទ្ធផលខុសគ្នាទាំងស្រុងត្រូវបានទទួលនៅក្នុង "មីក្រូកូស" ដែលមិនខុសពីគ្នា (តួនាទីរបស់វាត្រូវបានលេងដោយបំពង់សាកល្បងដែលមាន 0.5 សង់ទីម៉ែត្រ 3 នៃល្បាយសារធាតុចិញ្ចឹមដែលប៉ារ៉ាមិចត្រូវបានលាក់ដោយផ្នែក)។ ក្នុងករណីនេះលទ្ធផលគឺខុសគ្នា។ ពេលខ្លះសត្វមំសាសីបានស្លាប់ ហើយសត្វឈ្មោលក៏កើនឡើង។ ទោះយ៉ាងណាក៏ដោយ ប្រសិនបើចំនួនថ្មីនៃ ciliates ត្រូវបានណែនាំជាទៀងទាត់ទៅក្នុងមីក្រូកូសនោះ "រលកជីវិត" កើតឡើងតាមកាលកំណត់ កំឡុងពេលនោះការកើនឡើងនៃចំនួនសត្វឈ្មោលបណ្តាលឱ្យមានការកើនឡើងជាបន្តបន្ទាប់នៃចំនួនសត្វមំសាសី និងការថយចុះបរិមាណនៃសត្វទីមួយ។ បណ្តាលឱ្យមានការថយចុះនៃចំនួនប្រជាជន predator ។

ដូច្នេះលក្ខខណ្ឌបរិស្ថានប៉ះពាល់យ៉ាងខ្លាំងដល់លទ្ធផលនៃទំនាក់ទំនងដែលបានពិពណ៌នា។
ឥឡូវនេះសូមបន្តទៅលក្ខណៈសម្បត្តិរបស់សត្វមំសាសី។ ប្រសិនបើសត្វមំសាសីមានមធ្យោបាយវាយប្រហារដ៏ខ្លាំងក្លា (ដូចជា Didinium) នោះឥទ្ធិពលរបស់វាទៅលើចំនួនសត្វព្រៃគឺកាន់តែមុតស្រួច ហើយនៅក្នុងទឹកដីជាក់លាក់ណាមួយ សត្វមំសាសីនៅក្រោមលក្ខខណ្ឌមួយចំនួនអាចកំចាត់សត្វព្រៃទាំងស្រុង ឬបង្កើតការលើកទឹកចិត្តសម្រាប់សត្វព្រៃដើម្បីផ្លាស់ទី ( ប្រសិនបើវាមានលក្ខណៈ morphological សមស្រប) សមត្ថភាពរៀបចំសរីរវិទ្យា) ទៅកាន់ទីជម្រកផ្សេងទៀត។ បើទោះជាយ៉ាងណា សត្វមំាត្រូវបានការពារបានល្អ អាចទប់ទល់ រត់បានលឿន ឬបង្កើតឡើងវិញយ៉ាងខ្លាំងក្លា ហើយសត្វមំសាសីមានអាវុធវាយប្រហារខ្សោយ នោះបាតុភូតត្រូវបានកំណត់ចំពោះការប្រែប្រួលតាមកាលកំណត់ខាងលើ។ នៅក្នុងការកំណត់ធម្មជាតិ ទំនាក់ទំនងផ្សេងគ្នាអាចត្រូវបានគេសង្កេតឃើញ ហើយដូច្នេះជាមធ្យម តួនាទីរបស់សត្វមំសាសីមានសារសំខាន់ក្នុងការវិវត្តន៍។ ការពឹងផ្អែករបស់សត្វមំសាសី euryphage និង stenophage លើការប្រែប្រួលរបស់សត្វគឺខុសគ្នា។
សារៈសំខាន់ដ៏អស្ចារ្យ របៀបតឹងតែង. ឆ្នាំ ឬរយៈពេលនៃកង្វះអាហារូបត្ថម្ភកាត់បន្ថយយ៉ាងខ្លាំងនូវភាពធន់របស់បុគ្គលនៃប្រភេទសត្វដែលបានផ្តល់ឱ្យចំពោះកត្តាលុបបំបាត់ទាំងអស់ដែលបានរាយបញ្ជីខាងលើ។ ភាពអត់ឃ្លាននាំឱ្យមានការថយចុះនៃសកម្មភាព ការថយចុះនៃសភាវគតិការពារ ការចុះខ្សោយនៃភាពស៊ាំប្រឆាំងនឹងការឆ្លងមេរោគ ការថយចុះនៃការមានកូន។ ល នីមួយ, ភាពអត់ឃ្លានរបស់វាមិនលើសពី 5-10% ។ ក្នុងកំឡុងឆ្នាំទុរ្ភិក្ស ភាពរាំងស្ងួតឈានដល់ ២០-២៥% ចំនួនកូនកំប្រុកជាមធ្យម ១-៥ ចំនួនកំប្រុកក្មេង ២-៣ ក្បាល។ ក្នុងអំឡុងពេលជាច្រើនឆ្នាំនៃការបន្តពូជដ៏រឹងមាំនៃ lemmings ក្រោយមកទៀតនៅក្រោមឥទ្ធិពលនៃការខ្វះខាតអាហារបានប្រញាប់ប្រញាល់ក្នុងចំនួនដ៏ច្រើនទៅកាន់ទីជម្រកថ្មី។ សត្វជាច្រើនបានស្លាប់នៅពេលព្យាយាមជម្នះឧបសគ្គទឹក ហើយភាគច្រើនមកពីការវាយប្រហារដោយសត្វមំសាសី។ សត្វទីទុយប៉ូល កញ្ជ្រោង កញ្ជ្រោងអាក់ទិក និងសត្វដែលស្រេកឃ្លាន ប្រញាប់ប្រញាល់បន្ទាប់ពី lemmings សត្វរមាំង. បន្ទាប់ពីការវង្វេងបែបនេះចំនួនសត្វថយចុះយ៉ាងខ្លាំង។
ដូច្នេះ ប្រភេទសត្វនីមួយៗកំពុងជួបប្រទះសម្ពាធក្នុងការលុបបំបាត់ជានិច្ចពីកត្តាបរិស្ថាន biotic និង abiotic ។ កត្តាទាំងអស់ខាងលើធ្វើសកម្មភាពរួមគ្នាជាប្រព័ន្ធនៃកត្តា។ ពួកវាមួយចំនួនក្នុងឆ្នាំដែលបានផ្តល់ឱ្យគឺជិតនឹងកម្រិតល្អបំផុតសម្រាប់ប្រភេទសត្វដែលបានផ្តល់ឱ្យ ផ្ទុយទៅវិញ ពួកវាខ្លះមានផលប៉ះពាល់ក្នុងការលុបបំបាត់។ ការរួមបញ្ចូលគ្នានៃកត្តាជាក់លាក់ (ឧទាហរណ៍ សីតុណ្ហភាព និងសំណើម) ក៏មានឥទ្ធិពលយ៉ាងខ្លាំងលើរាងកាយផងដែរ។ តាមក្បួនវាគឺជាការរួមបញ្ចូលគ្នានៃកត្តាបរិស្ថានផ្សេងៗដែលមានឥទ្ធិពល។
សក្តានុពលសម្រាប់ការរស់រានមានជីវិតនៅក្នុងលក្ខខណ្ឌទាំងនេះត្រូវបានកំណត់ដោយហេតុផលពីរ។ ទីមួយវាអាស្រ័យលើស្ថានភាពនៃកត្តានាំមុខសម្រាប់ប្រភេទនេះ។ ឧទាហរណ៍ ប្រសិនបើសម្រាប់ប្រភេទសត្វដែលបានផ្តល់ឱ្យ សីតុណ្ហភាព និងសំណើមមានសារៈសំខាន់បំផុត ហើយស្ថានភាពនៃកត្តាទាំងនេះគឺល្អប្រសើរបំផុត នោះភាពមិនអំណោយផលដែលគេស្គាល់នៃកត្តាផ្សេងទៀតនឹងមានផលប៉ះពាល់តិចជាងទៅលើចំនួនប្រជាជននៃប្រភេទសត្វ។
ទោះជាយ៉ាងណាក៏ដោយ កម្រិតនៃភាពធន់នៃប្រភេទសត្វក្នុងការលុបបំបាត់កត្តាបរិស្ថានគឺមានសារៈសំខាន់ជាការសម្រេចចិត្ត។ ភាពធន់នៃប្រភេទសត្វត្រូវបានកំណត់ដោយភាពធន់នៃអេកូឡូស៊ីរបស់វា ដែលសំដៅលើវិសាលភាពនៃសមត្ថភាពរបស់វាក្នុងការសម្របខ្លួនទៅនឹងការផ្លាស់ប្តូរលក្ខខណ្ឌបរិស្ថាន។ Valence អាចមានលក្ខណៈទូលំទូលាយ ហើយប្រភេទសត្វបែបនេះត្រូវបានគេហៅថា euryadaptive ឬតូចចង្អៀត (ប្រភេទសត្វ stenadaptive) ។ ទោះយ៉ាងណាក៏ដោយ មិនថា valence មានទំហំប៉ុនណាក៏ដោយ វាមិនដែលស្មើរនឹងកត្តាលុបបំបាត់ទាំងអស់នោះទេ។ ជាឧទាហរណ៍ ប្រភេទសត្វមួយអាចមានភាពប្រែប្រួលនៃប្រព័ន្ធអេកូឡូស៊ីយ៉ាងទូលំទូលាយទាក់ទងនឹងការប្រែប្រួលសីតុណ្ហភាព (ប្រភេទសត្វ eurythermal) ប៉ុន្តែត្រូវមានឯកទេសខ្ពស់ទាក់ទងនឹងរបបនៃការផ្តល់អាហារ (stenophages) ឬមានលក្ខណៈ stenothermic ប៉ុន្តែក្នុងពេលតែមួយ euryphage ជាដើម។ លើសពីនេះទៀត Euryadaptivity មានដែនកំណត់របស់វា។ ឧទាហរណ៍ pasyuk គឺជាឧទាហរណ៍ធម្មតានៃទម្រង់ euryadaptive ទោះយ៉ាងណាក៏ដោយ ដូចដែលយើងបានឃើញ ភាពប្រែប្រួលនៃអេកូឡូស៊ីរបស់វាមានដែនកំណត់ជាក់លាក់របស់វា។
ក្នុងករណីណាក៏ដោយ កម្រិតនៃ euryadaptation ទាក់ទងទៅនឹងកត្តាបរិស្ថានដែលបានផ្តល់ឱ្យ និងកត្តាទាំងអស់នៃស្ថានីយ៍ និង biocenosis ទាំងមូល គឺជាមូលដ្ឋានសម្រាប់កំណត់លក្ខណៈសក្តានុពលនៃការរស់រានមានជីវិតនៃប្រភេទសត្វ ហើយសក្តានុពលនៃការរស់រានមានជីវិតជាមធ្យមគឺ សមាមាត្រដោយផ្ទាល់ទៅនឹង valence អេកូឡូស៊ីនៃប្រភេទសត្វ។
ចូរយើងផ្តល់ឧទាហរណ៍ខ្លះៗ។ ក្នុងរយៈពេលជាច្រើនឆ្នាំជាមួយនឹងរបបផ្តល់អាហារថយចុះ សក្តានុពលនៃការរស់រានមានជីវិតរបស់ euryphages គឺខ្ពស់ជាង stenophages ។ សត្វមំសាសីមួយចំនួននៅពេលដែលខ្វះអាហារមួយប្រភេទ ប្តូរទៅមួយទៀត ដែលអនុញ្ញាតឱ្យវាជៀសផុតពីស្ថានភាពលំបាក។ ធម្មជាតិ omnivorous នៃ ចំនួន នៃ ប្រភេទ សត្វល្អិត អនុញ្ញាត ឱ្យ ពួក គេ ដើម្បី រស់ នៅ ក្នុង អវត្ដមាន នៃ រុក្ខជាតិ ជាក់លាក់ ។ Stenophages ស្លាប់នៅក្រោមលក្ខខណ្ឌទាំងនេះ។ ដូច្នេះឧទាហរណ៍ការប្រយុទ្ធប្រឆាំងនឹងសត្វល្អិតដែលបង្កគ្រោះថ្នាក់ឬ nematodes - euryphages ជាក្បួនគឺពិបាកជាងជាមួយ stenophages ។
ដូច្នេះ សក្តានុពលជីវសាស្រ្តនៃប្រភេទសត្វ ភាពរឹងមាំរបស់វា គឺជាលទ្ធផលជាក់លាក់នៃបរិមាណពីរ - សក្តានុពលបន្តពូជ និងសក្តានុពលនៃការរស់រានមានជីវិត ដែលនៅក្នុងវេនត្រូវបានកំណត់ដោយកម្រិតនៃ valence អេកូឡូស៊ីនៃប្រភេទសត្វ។ នៅក្រោមឥទ្ធិពលនៃការរួមបញ្ចូលគ្នានៃកត្តាលុបបំបាត់ដែលបានរៀបរាប់ខាងលើចំនួននៃមនុស្សពេញវ័យនៃជំនាន់មួយគឺតែងតែតិចជាងចំនួនទារកទើបនឹងកើត។ ការពិតនេះត្រូវបានសិក្សាយ៉ាងល្អតាមរយៈការវិភាគបរិមាណនៃសក្ដានុពលនៃចំនួនកូនដែលកើតក្នុងឆ្នាំដែលបានផ្តល់ឱ្យ និងរបស់ពួកគេ វាសនាអនាគត. តាមក្បួនមួយ (ដូចដែលលោក Darwin បានចង្អុលបង្ហាញ) អត្រាមរណភាពខ្ពស់ត្រូវបានគេសង្កេតឃើញក្នុងចំណោមបុគ្គលវ័យក្មេង ដែលនាំឱ្យមានការថយចុះយ៉ាងឆាប់រហ័សនៃចំនួនកូនចៅ។ ការវិភាគសមាសភាពនៃចំនួនប្រជាជនតាមអាយុ និងគណនាភាគរយនៃប្រភេទនីមួយៗ ក្រុមអាយុចំពោះចំនួនសរុបនៃបុគ្គល (នេះអាចធ្វើបាន ជាពិសេសទាក់ទងនឹងហ្គេមសត្វ និងបក្សី) វាអាចត្រូវបានបង្កើតឡើងថាការថយចុះនៃចំនួនតែងតែគោរពតាមខ្សែកោងជាក់លាក់មួយ។ ជាឧទាហរណ៍ តួរលេខបង្ហាញពីការថយចុះនៃចំនួនកូនកំប្រុក។ ដូចដែលអាចមើលឃើញនៅឆ្នាំដំបូងនៃអត្រាមរណៈនៃជីវិតគឺខ្ពស់ បន្ទាប់មកអត្រារបស់វាធ្លាក់ចុះ ហើយការស្លាប់នៃទម្រង់មនុស្សពេញវ័យកាន់តែតិច។

ខ្សែកោងស្រដៀងគ្នានេះប្រហែលជាត្រូវបានគូររួចហើយសម្រាប់ប្រភេទសត្វមួយចំនួនធំ។ តួលេខដូចគ្នាបង្ហាញពីសក្ដានុពលនៃចំនួនអាយុ spruce ។ វាងាយស្រួលក្នុងការមើលឃើញភាពស្រដៀងគ្នានៃខ្សែកោងទាំងនេះ ទោះបីជាមានភាពខុសគ្នាយ៉ាងជ្រាលជ្រៅរវាងវត្ថុជីវសាស្រ្ត (កំប្រុក និង spruce) ក៏ដោយ។ វាច្បាស់ណាស់ថាយើងកំពុងដោះស្រាយនៅទីនេះជាមួយនឹងបុព្វហេតុទូទៅមួយ។ ចុងក្រោយនេះគឺជាការតស៊ូដើម្បីអត្ថិភាព ដែលវត្ថុជីវសាស្រ្តទាំងអស់មានប្រធានបទស្មើគ្នា។ ខ្សែកោងបង្ហាញថាការតស៊ូដើម្បីអត្ថិភាពមានសារៈសំខាន់ក្នុងការលុបបំបាត់ទាំងស្រុង: បុគ្គលខ្លះស្លាប់។ ដូច្នេះ ការតស៊ូដើម្បីអត្ថិភាព គឺជាកត្តាលុបបំបាត់ធម្មជាតិ ដែលកំណត់ការរលត់នៃសមុដ្ឋានតិច និងការរស់រានមានជីវិតដែលសមជាង។
ប្រភេទនៃការលុបបំបាត់
វាមានសារៈសំខាន់ណាស់ក្នុងការស្វែងយល់ថាតើសារៈសំខាន់នៃការវិវត្តនៃសកម្មភាពលុបបំបាត់ការតស៊ូដើម្បីអត្ថិភាពគឺជាអ្វី។ ប្រសិនបើបុគ្គលខ្លះស្លាប់ និងខ្លះទៀតនៅរស់ នោះសំណួរកើតឡើងថាតើអ្វីកំណត់ភាពខុសគ្នានេះ?
ចម្លើយចំពោះសំណួរនេះនឹងកាន់តែច្បាស់ប្រសិនបើយើងពិចារណាអំពីលក្ខណៈនៃការលុបបំបាត់ ប្រភេទរបស់វា ដែលយើងនឹងពិចារណាឥឡូវនេះ។
ក) ការលុបបំបាត់ដោយមិនជ្រើសរើសបុគ្គល (ចៃដន្យ) ទាក់ទងនឹងបុគ្គល. ឧទាហរណ៍ដូចជាការស្លាប់របស់សត្វលេបនៅលើបន្លាដ៏រឹងមាំនៃ burdock មួយ។ ការស្លាប់នេះគឺចៃដន្យ ហើយកម្រត្រូវបានគេសង្កេតឃើញ (ករណីស្រដៀងគ្នានេះត្រូវបានពិពណ៌នាសម្រាប់សត្វប្រចៀវមួយ)។ ទោះយ៉ាងណាក៏ដោយ មានករណីបែបនេះច្រើនណាស់នៅក្នុងជីវិតរបស់រុក្ខជាតិ និងសត្វ ហើយពួកវាអាចមានសារៈសំខាន់ជាក់លាក់មួយ ឧទាហរណ៍ ក្នុងអំឡុងពេលដាក់សំបុក នៅពេលដែលការស្លាប់ដោយចៃដន្យនៃស្ត្រីដែលបំបៅកូនដោយអចេតនា នាំឱ្យមានការស្លាប់របស់កូនចៅទាំងអស់។ តាមទ្រឹស្ដី មនុស្សម្នាក់អាចស្រមៃថា សត្វមូសណាមួយ ហើយដូច្នេះកូនចៅរបស់គាត់ អាចស្លាប់តាមរបៀបនេះ។

ខ) ការលុបបំបាត់ក្រុមដែលមិនជ្រើសរើស (ចៃដន្យ)ការព្រួយបារម្ភលែងជាបុគ្គលម្នាក់ៗទៀតហើយ ប៉ុន្តែក្រុមបុគ្គល និងត្រូវបានកំណត់ដោយឥទ្ធិពលរីករាលដាលនៃកត្តាបំផ្លិចបំផ្លាញដោយចៃដន្យមួយចំនួន ឧទាហរណ៍ ភ្លើងឆេះព្រៃមានកម្រិត ទឹកជំនន់រដូវរងាក្នុងតំបន់ ការដួលរលំភ្នំ សាយសត្វក្នុងស្រុកភ្លាមៗ (ជាពិសេសបន្ទាប់ពីភ្លៀង) ការលាង ឃ្លាតឆ្ងាយពីសត្វ ឬរុក្ខជាតិដោយទឹកភ្លៀង ជាដើម ។ល។ ក្នុងករណីបែបនេះ ទាំង "ប្រែប្រួល" និង "មិនសម្របខ្លួន" ស្លាប់។ ក្នុងករណីនេះ ការស្លាប់អាចប៉ះពាល់ដល់ក្រុមបុគ្គលនៃសមាសភាពហ្សែនជាក់លាក់មួយ។ ឧទាហរណ៍ ប្រសិនបើ mutant មួយបានកើតឡើងដែលមិនមានពេលវេលាក្នុងការបន្តពូជក្នុងបរិមាណច្រើន រីករាលដាលយឺត និងមានតំបន់តូចមួយ (កណ្តាល) នៃការចែកចាយ នោះការលុបបំបាត់ក្រុមដោយចៃដន្យអាចគ្របដណ្តប់សមាសភាពបុគ្គលទាំងមូលនៃកូនចៅរបស់ mutant ។ ដោយមិនគិតពីអត្ថប្រយោជន៍ ឬគ្រោះថ្នាក់នៃការផ្លាស់ប្តូរដែលបានផ្តល់ឱ្យ អ្នកដឹកជញ្ជូនទាំងអស់របស់វាអាចត្រូវបានបំផ្លាញ។ ដូច្នេះ ការលុបបំបាត់ក្រុមដោយចៃដន្យនៅក្នុងករណីបែបនេះអាចប៉ះពាល់ដល់សមាសភាពហ្សែននៃប្រភេទសត្វ ទោះបីជាវានៅតែមិនមានសារៈសំខាន់ក្នុងការវិវត្តន៍ឈានមុខគេក៏ដោយ។
វី) ការលុបបំបាត់មហន្តរាយដែលមិនរើសអើងកើតឡើងជាមួយនឹងការចែកចាយកាន់តែទូលំទូលាយនៃកត្តាបំផ្លិចបំផ្លាញ ឧទាហរណ៍ សាយសត្វមិនធម្មតា ទឹកជំនន់ ភ្លើងឆេះព្រៃដែលបានលេបត្របាក់តំបន់ធំ គ្រោះរាំងស្ងួតពិសេស លំហូរកម្អែ និងគ្រោះមហន្តរាយផ្សេងទៀតដែលរាលដាលពាសពេញតំបន់ដ៏ធំ។ ហើយក្នុងករណីនេះ ទាំង "សម្របខ្លួន" និង "មិនសម្របខ្លួន" ត្រូវវិនាស។ ទោះជាយ៉ាងណាក៏ដោយ ទម្រង់នៃការលុបបំបាត់នេះអាចមានសារៈសំខាន់ក្នុងការវិវត្តន៍ដ៏អស្ចារ្យ ដែលជះឥទ្ធិពលដល់សមាសភាពហ្សែននៃប្រភេទសត្វកាន់តែមានប្រសិទ្ធភាព និងមានឥទ្ធិពលខ្លាំងលើ biocenoses ទាំងមូល។
Naumov (1939) បានសង្កេតឃើញថាជាលទ្ធផលនៃទឹកភ្លៀងនៅតំបន់ steppe នៃភាគខាងត្បូងអ៊ុយក្រែន burrows rodent ត្រូវបានជន់លិចដែលបណ្តាលឱ្យមានការថយចុះយ៉ាងខ្លាំងនៃចំនួន voles ។ ទន្ទឹមនឹងនេះដែរប្រជាជនក្នុងស្រុកនៃកណ្តុរ Kurgan មិនមានការផ្លាស់ប្តូរគួរឱ្យកត់សម្គាល់ទេ។ នេះត្រូវបានពន្យល់ដោយការចល័តកាន់តែច្រើនរបស់សត្វកណ្តុរបើប្រៀបធៀបទៅនឹង voles ។ នៅពេលដែលព្រិលរលាយនៅនិទាឃរដូវ ប្រហោងសត្វកកេរត្រូវបានបិទដោយដោតទឹកកក ហើយសត្វកណ្ដុរស្លាប់ដោយការអត់ឃ្លាន ខណៈដែលសត្វកណ្តុររស់រានមានជីវិតដោយសារតែវាស្តុកអាហារនៅក្នុងបន្ទប់ក្រោមដី។ ឧទាហរណ៍ដែលបានជ្រើសរើសបង្ហាញពីឥទ្ធិពលនៃវិសមភាពជីវសាស្រ្តនៃប្រភេទសត្វពីរទាក់ទងនឹងកត្តាខាងក្រៅដូចគ្នា។ វាច្បាស់ណាស់ថាទំនាក់ទំនងបែបនេះអាចបណ្តាលឱ្យមានការវិវត្តន៍នៃ biocenoses (ជោគជ័យ) និងការផ្លាស់ប្តូរនៅក្នុងសមាសភាពប្រភេទសត្វនៃប្រភេទបុគ្គល គ្រួសារ។ល។
ឧទាហរណ៏នៃការលុបបំបាត់គ្រោះមហន្តរាយគឺការស្លាប់ដ៏ធំនៃ muskrats ក្នុងអំឡុងពេលទឹកជំនន់រដូវរងារឬការស្លាប់នៃ partridges ពណ៌ប្រផេះ។ រដូវរងាដ៏អាក្រក់ 1839/40 ជាដើម។ សញ្ញាសំខាន់នៃការលុបបំបាត់គ្រោះមហន្តរាយគឺការបំផ្លិចបំផ្លាញដ៏ធំនៃបុគ្គលនៃប្រភេទសត្វ ដោយមិនគិតពីសក្តានុពលនៃការរស់រានមានជីវិតរបស់ពួកគេ។
ឆ) ការលុបបំបាត់សរុប (ទូទៅ). វាក៏មានតម្លៃផងដែរក្នុងការគូសបញ្ជាក់ទម្រង់នៃការលុបបំបាត់នេះ ក្រោមការដែលចំនួនប្រជាជនទាំងមូលនៃប្រភេទសត្វស្លាប់ ពោលគឺ បុគ្គលទាំងអស់ដែលរួមបញ្ចូលនៅក្នុងសមាសភាពរបស់វា។ ទម្រង់នៃការលុបបំបាត់នេះក៏មិនរើសអើងដែរ។ វាអាចទៅរួចក្នុងករណីដែលជួរនៃប្រភេទសត្វមានទំហំតូច ឬនៅពេលក្រោយត្រូវបានប៉ះពាល់ទាំងស្រុងដោយកត្តាមិនអំណោយផលមួយចំនួន។ ជាឧទាហរណ៍ ប្រហែលជាការលុបបំបាត់ទាំងស្រុងគឺជាមូលហេតុនៃការស្លាប់របស់ mammoth នៅស៊ីបេរី។ វាជាការងាយស្រួលក្នុងការស្រមៃថាការលុបបំបាត់ទាំងស្រុងអាចនាំទៅដល់ការស្លាប់របស់ប្រជាជនទាំងមូលនៃប្រភេទសត្វឆ្លងមួយចំនួន ដោយកាន់កាប់ឧទាហរណ៍ កំពូលភ្នំមួយ ឬកោះតូចមួយ ដែលគ្របដណ្តប់ទាំងស្រុងដោយមួយចំនួន។ គ្រោះធម្មជាតិលល។
តាមអ្វីដែលបាននិយាយទាក់ទងនឹងការលុបបំបាត់ទាំងស្រុង វាច្បាស់ណាស់ថា ការបែងចែកដាច់ខាតរវាងទម្រង់នៃការលុបបំបាត់ដោយមិនរើសអើងក្នុងបញ្ជីគឺមិនអាចទៅរួចទេ។ ភាគច្រើនត្រូវបានកំណត់ ដូចដែលយើងឃើញដោយទំហំនៃប្រភេទសត្វ ចំនួនបុគ្គលដែលរួមបញ្ចូលនៅក្នុងសមាសភាពរបស់វា។ ការលុបបំបាត់ ដែលមានសារៈសំខាន់ជាក្រុមសម្រាប់ប្រភេទសត្វមួយចំនួន នឹងមានសរុបសម្រាប់ប្រភេទសត្វដទៃទៀត។ ភាគច្រើនត្រូវបានកំណត់ដោយលក្ខណៈសម្បត្តិនៃទម្រង់រស់នៅទាំងនោះដែលត្រូវបានប៉ះពាល់ទៅនឹងកត្តាលុបបំបាត់ទាំងនេះ។ ជាឧទាហរណ៍ ភ្លើងឆេះព្រៃដែលមានកម្រិតនឹងបំផ្លាញរុក្ខជាតិ ខណៈសត្វអាចគេចចេញពីវាបាន។ ទោះជាយ៉ាងណាក៏ដោយចំនួនសត្វគឺមិនស្មើគ្នាក្នុងរឿងនេះ។ ទម្រង់ដីតូចខ្លាំងដែលរស់នៅក្នុងព្រៃនឹងងាប់ជាច្រើន។ រឿងដដែលនឹងកើតឡើងចំពោះសត្វល្អិតជាច្រើន ឧទាហរណ៍ ស្រមោចព្រៃ សត្វល្អិតជាច្រើនជាដើម ។ កង្កែបស្មៅ, ជីងចក់ viviparous ជាដើម - ជាទូទៅ ទម្រង់ទាំងអស់ដែលល្បឿនដកថយគឺទាបជាងល្បឿននៃការរីករាលដាលនៃភ្លើង។ ថនិកសត្វ និងបក្សីនឹងអាចចាកចេញបានក្នុងករណីភាគច្រើន។ ទោះជាយ៉ាងណាក៏ដោយនៅទីនេះច្រើនពេកត្រូវបានកំណត់ដោយដំណាក់កាលនៃការអភិវឌ្ឍន៍បុគ្គល។ វានឹងមិនមានភាពខុសគ្នាច្រើនរវាងពងសត្វឃ្មុំ និងស៊ុតបក្សី មេអំបៅ និងកូនមាន់ទេ។ ជាការពិតណាស់ក្នុងគ្រប់ករណីទាំងអស់ ទម្រង់ដែលរងគ្រោះច្រើនបំផុតគឺជាទម្រង់ដែលមានទីតាំង ដំណាក់កាលដំបូងការអភិវឌ្ឍន៍បុគ្គល។
ឃ) ការលុបបំបាត់ការជ្រើសរើសមានសារៈសំខាន់នៃការវិវត្តន៍ដ៏អស្ចារ្យបំផុត ចាប់តាំងពីក្នុងករណីនេះ ឥទ្ធិពលចម្បងនៃការតស៊ូដើម្បីអត្ថិភាពត្រូវបានធានា ពោលគឺការស្លាប់របស់អ្នកដែលសមតិចបំផុត និងការរស់រានមានជីវិតរបស់អ្នកសមបំផុត។ ការលុបបំបាត់ដោយជ្រើសរើសគឺផ្អែកលើតំណពូជហ្សែនរបស់បុគ្គល ឬក្រុមរបស់ពួកគេ ហើយដូច្នេះនៅលើធម្មជាតិនៃការកែប្រែ និងលទ្ធផលវិសមភាពជីវសាស្រ្តនៃទម្រង់ផ្សេងៗ។ វាគឺនៅក្នុងករណីនេះដែលការកែលម្អធម្មជាតិ និងការវិវត្តន៍រីកចម្រើននៃប្រភេទសត្វកើតឡើង។
ការតស៊ូក្នុងភាពជាក់លាក់ និងអន្តរជាក់លាក់សម្រាប់អត្ថិភាព
ការលុបបំបាត់ដោយជ្រើសរើសគឺជាពេលលក្ខណៈពិសេសបំផុតនៃការតស៊ូដើម្បីអត្ថិភាព ដែលជាការបញ្ចេញមតិពិតប្រាកដរបស់វា។ តាមរយៈការលុបទម្រង់ដែលមិនពេញចិត្តដោយជ្រើសរើស ការរក្សាទុកសំណល់នៃបុគ្គល ឬក្រុមបុគ្គលដែលសម្របខ្លួនបំផុតត្រូវបានសម្រេច។
សំណួរកើតឡើង តើក្នុងក្រុមជាក់លាក់ណាមួយនៃបុគ្គលដែលការដកចេញដោយជ្រើសរើសមានសារសំខាន់នៃការវិវត្តន៍ដ៏អស្ចារ្យបំផុត? ដាវីនបានចង្អុលបង្ហាញថា សំណួរនេះទាក់ទងនឹងសំណួរនៃការតស៊ូដើម្បីអត្ថិភាព។ គាត់បានភ្ជាប់សារៈសំខាន់ដ៏ធំបំផុតចំពោះការតស៊ូ intraspecific សម្រាប់អត្ថិភាព។ ការប្រកួតប្រជែងខ្លាំងបំផុតរវាងទម្រង់នានាកើតឡើងនៅក្នុងប្រភេទសត្វដូចគ្នា ចាប់តាំងពីតម្រូវការរបស់បុគ្គលដែលមានប្រភេទដូចគ្នាគឺនៅជិតគ្នាបំផុត ដូច្នេះហើយការប្រកួតប្រជែងរវាងពួកវាគឺខ្លាំងជាង។
យើងដឹងរួចមកហើយថា បុគ្គលដែលមានប្រភេទដូចគ្នាមានជីវសាស្រ្តមិនស្មើគ្នា ពោលគឺពួកគេមានឱកាសខុសៗគ្នាក្នុងការទប់ទល់នឹងកត្តាបរិស្ថានដែលបំផ្លិចបំផ្លាញ។ វិសមភាពជីវសាស្រ្តនេះត្រូវបានបញ្ជាក់យ៉ាងច្បាស់នៅក្នុងការពិតដែលថាបុគ្គលផ្សេងគ្នាមានភាពខុសគ្នាមួយចំនួននៅក្នុងសក្តានុពលជីវសាស្រ្ត។
លើសពីនេះ យើងដឹងថាមានការប្រកួតប្រជែងដោយប្រយោល និងដោយផ្ទាល់រវាងបុគ្គលម្នាក់ៗ ហើយវា (យោងទៅតាមលោក Darwin) គឺកាន់តែខ្លាំងក្លា បុគ្គលដែលប្រកួតប្រជែងកាន់តែជិតស្និទ្ធនឹងគ្នាទៅវិញទៅមកនៅក្នុងតម្រូវការរបស់ពួកគេ។ ពីទីនេះ វាច្បាស់ណាស់ថា បុគ្គលនីមួយៗនៃប្រភេទសត្វមាន "បន្ទុក" សំខាន់ទ្វេរដង៖ ក) វាទប់ទល់នឹងវិសាលភាពនៃសក្តានុពលជីវសាស្ត្ររបស់វា លុបបំបាត់កត្តាបរិស្ថាន និង ខ) ប្រកួតប្រជែងជាចម្បងសម្រាប់អាហារ និងលំហ។ បុគ្គលផ្សេងទៀតនៃប្រភេទ។ វាច្បាស់ដូចគ្នាថាការតស៊ូជាមួយកត្តាលុបបំបាត់គឺកាន់តែខ្លាំង ការប្រកួតប្រជែងកាន់តែខ្លាំងជាមួយបុគ្គលដទៃទៀតនៃប្រភេទសត្វ។ យ៉ាងណាមិញ ការប្រកួតប្រជែងនេះគឺដូចជា "បន្ទុកបន្ថែម" ដែលធ្វើអោយការតស៊ូដើម្បីអត្ថិភាពកាន់តែធ្ងន់ធ្ងរឡើង។ ពីអ្វីដែលបានរៀបរាប់ខាងលើ វាច្បាស់ណាស់ថាការតស៊ូសរុបសម្រាប់អត្ថិភាពគឺខ្លាំងជាពិសេសរវាងបុគ្គលដែលមានផលប្រយោជន៍ជីវិតស្រដៀងគ្នា ពោលគឺបុគ្គលដែលមានលក្ខណៈអេកូឡូស៊ីដូចគ្នា។
ទីផ្សារពិសេសមួយត្រូវបានយល់ថាជាភាពស្មុគស្មាញនៃលក្ខខណ្ឌបរិស្ថានសម្ភារៈដែលបុគ្គល ក) សម្របខ្លួនបានច្រើនបំផុត ខ) ទាញយកធនធានអាហារ និង គ) មានឱកាសបង្កើតឡើងវិញយ៉ាងខ្លាំងក្លាបំផុត។ កាន់តែច្បាស់ជាងនេះទៅទៀត ទីផ្សារពិសេសគឺជាភាពស្មុគស្មាញនៃលក្ខខណ្ឌបរិស្ថានសម្ភារៈដែលសក្តានុពលជីវសាស្រ្តនៃប្រភេទសត្វមានការបញ្ចេញមតិពេញលេញបំផុត។
ឧទាហរណ៍សម្រាប់កំហុសក្រហម ទីផ្សារពិសេសរបស់វាគឺដី។ សត្វល្អិតស៊ីចំណីលើសាកសពរបស់សត្វល្អិត ដោយបឺតទឹកចេញ ដោយមានជំនួយពី proboscis របស់វា។ ដីបម្រើជាប្រភពសំណើមសម្រាប់វា។ អ្នកនិពន្ធបានសង្កេតឃើញជាញឹកញាប់ថា សត្វល្អិតក្រហមបានទម្លាក់ proboscis របស់វាទៅក្នុងដី ហើយបឺតទឹក។ គម្របបន្លែបម្រើជាជម្រកសម្រាប់វា។ ការបន្តពូជក៏កើតឡើងនៅលើផែនដីដែរ។ ស្រីបង្កើតរណ្ដៅតូចៗនៅក្នុងដីដែលពងរបស់ពួកគេត្រូវបានដាក់។ ការភ្ជាប់ទៅនឹងដីជាទីផ្សារពិសេសក៏បណ្តាលឱ្យមានការផ្លាស់ប្តូរនៅក្នុងការរៀបចំនៃកំហុសក្រហម។ ស្លាបខាងក្រោយ (ជើងហោះហើរ) របស់វាត្រូវបានប្រែក្លាយទៅជាកំណាត់។ អាស្រ័យហេតុនេះ ការភ្ជាប់ទៅនឹងដីនាំឱ្យបាត់បង់សមត្ថភាពហោះហើរ។ ឧទាហរណ៍ដ៏ល្អមួយទៀតគឺ ទីផ្សារពិសេសរបស់ muskrat ។ តម្រូវការសំខាន់ៗទាំងអស់របស់វា ហើយសំខាន់ជាងនេះទៅទៀត អាហាររូបត្ថម្ភដ៏បរិបូរណ៍ចាំបាច់គឺត្រូវបានពេញចិត្តនៅតំបន់ទំនាបលិចទឹក និងទន្លេខាងក្រោយ។ វាគួរឱ្យកត់សម្គាល់ថាការបន្តពូជក៏ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងធាតុទឹកផងដែរ។ អ្នកនិពន្ធបានសង្កេតម្តងហើយម្តងទៀតនូវ "ល្បែង" នៃ muskrats នៅក្នុងទឹកហើយនៅក្នុង vivarium ដែលត្រូវបានសាងសង់ជាពិសេសការប៉ុនប៉ងនៅ coitus ធ្វើឡើងនៅក្នុងទឹក (Paramonov, 1932) ។ ដូច្នេះ បរិមាណទឹកនៃបឹងលិចទឹក និងទឹកខាងក្រោយ ដែលសម្បូរទៅដោយបន្លែ និងធនធានអាហារផ្សេងទៀត ក្លាយជាទីផ្សារពិសេសរបស់ muskrat ដែលវាត្រូវបានសម្របតាមលក្ខណៈឈានមុខគេទាំងអស់នៃអង្គការ morphophysiological របស់វា។ នោះហើយជាមូលហេតុដែល muskrat burrows ជាក្បួនមានច្រកចេញតែមួយគត់ - ចូលទៅក្នុងទឹក។
ចាប់តាំងពីបុគ្គលនៃប្រភេទដូចគ្នា ជាក្បួនត្រូវបានកំណត់ដោយ niches ដូចគ្នា ឬមានគុណភាព វាគឺជាការតស៊ូ intraspecific សម្រាប់អត្ថិភាពដែលខ្លាំងបំផុត។ ដូច្នេះ ដាវីនបានកំណត់យ៉ាងត្រឹមត្រូវនូវការតស៊ូក្នុងអន្តរជាក់លាក់ថាជាប្រភេទឯករាជ្យនៃទំនាក់ទំនងប្រកួតប្រជែងរវាងសារពាង្គកាយ។ ចូរយើងពិចារណាឧទាហរណ៍មួយចំនួននៃការតស៊ូមិនជាក់លាក់សម្រាប់អត្ថិភាព ដែលបង្កើតឡើងដោយការសង្កេតតាមវាល និងការសិក្សាពិសោធន៍។ ចូរយើងរំលឹកឡើងវិញនូវទំនាក់ទំនងរវាងកញ្ជ្រោងពណ៌ស និងពណ៌ខៀវដែលបានពិពណ៌នាខាងលើ (ការតស៊ូដោយប្រយោលសម្រាប់អត្ថិភាព)។ នៅក្នុងលក្ខខណ្ឌនៃ tundra ដីគោក កញ្ជ្រោងអាកទិកពណ៌សគ្របដណ្តប់ ហើយនៅក្នុងលក្ខខណ្ឌនៃកោះ Commander ពណ៌ខៀវគ្របដណ្តប់។ ឧទាហរណ៍មួយទៀតគឺទំនាក់ទំនងរវាងទម្រង់ធម្មតា និង melanistic នៃមេអំបៅខែ birch ។ ទម្រង់ស្លាបស្រាលធម្មតា (Amphidasis betularia) បានគ្រប់គ្រងដំបូង ប៉ុន្តែក្នុងទសវត្សរ៍ទី 60 នៅប្រទេសអង់គ្លេស (នៅតំបន់ជុំវិញទីក្រុង Manchester) ទម្រង់ស្លាបងងឹត (A. b. doubledayria) បានចាប់ផ្តើមបង្កើតឡើងវិញយ៉ាងសកម្ម។ ក្រោយមកទៀតបានជំនួសធម្មតា (ស្លាបស្រាល) ដំបូងបង្អស់នៅក្នុងប្រទេសអង់គ្លេស ហើយបន្ទាប់មក (ក្នុងទសវត្សរ៍ទី 80) ដំណើរការដូចគ្នានេះបានរីករាលដាលនៅក្នុង អឺរ៉ុបខាងលិច. Dementyev (1940) សំដៅទៅលើឧទាហរណ៍ខាងក្រោម។ សត្វពពែពណ៌ខៀវ (Anser coerulescens) ត្រូវបានជំនួសដោយសត្វ mutant ពណ៌សនៅក្នុងជួរភាគច្រើនរបស់វា។ នៅលើកោះ St. Vincent (ក្រុម Antilles នៃកោះ) ដែលជាមេឡានីននិយមនៃសត្វស្លាប Coereba saccharina បានកើតឡើង។ នៅឆ្នាំ 1878 សត្វ mutant បានក្លាយជាលេខលេចធ្លោ នៅឆ្នាំ 1903 ទម្រង់ធម្មតាត្រូវបានរកឃើញនៅក្នុងច្បាប់ចម្លងតែមួយ។ល។
ទិន្នន័យពិសោធន៍ក៏បញ្ជាក់ពីវត្តមាននៃការតស៊ូមិនជាក់លាក់សម្រាប់អត្ថិភាព។ ឧទាហរណ៍មួយគឺការសិក្សាដ៏ល្អឥតខ្ចោះរបស់ Sukachev (1923) ស្តីពីការស្លាប់នៃទម្រង់ហ្សែន intraspecific ផ្សេងៗនៃ dandelion ទូទៅ (Taraxacum officinale) ។ នៅលើដីឡូត៍ ផ្ការំដួលត្រូវបានគេសាបព្រោះក្នុងទម្រង់តំណពូជចំនួនបី ដែលកំណត់តាមធម្មតា A, B និង C។ ដំណាំត្រូវបានលាយបញ្ចូលគ្នា និងសុទ្ធ ក្រោមលក្ខខណ្ឌនៃការដាំមិនសូវល្អ និងក្រាស់។ អត្រាមរណៈក្នុងលក្ខខណ្ឌផ្សេងៗត្រូវបានពិនិត្យ ដូចដែលអាចមើលឃើញក្នុងតារាង។
សូមក្រឡេកមើលទិន្នន័យពីតារាងទាំងនេះ។

តារាងបង្ហាញថាទម្រង់ intraspecific ផ្សេងគ្នាត្រូវបានបែងចែកដោយគោរពតាមសក្តានុពលនៃការរស់រានមានជីវិតរបស់ពួកគេ។ លើសពីនេះទៅទៀតវាត្រូវបានបញ្ជាក់នៅទីនេះថាសក្តានុពលនៃការរស់រានមានជីវិតក៏ផ្លាស់ប្តូរនៅក្រោមលក្ខខណ្ឌផ្សេងៗគ្នា។ ដូច្នេះនៅក្នុងវប្បធម៌ដ៏កម្រមួយ អត្រាមរណៈកើនឡើងតាមលំដាប់ C-A-B នៅក្នុងវប្បធម៌សុទ្ធដ៏ក្រាស់ - B-A-C នៅក្នុងវប្បធម៌ចម្រុះដ៏កម្រ និងក្នុងវប្បធម៌ចម្រុះដ៏ក្រាស់ C-A-B ។

តារាងបង្ហាញថាទម្រង់ A, B និង C មានសក្តានុពលនៃការបន្តពូជខុសៗគ្នា។ អាស្រ័យហេតុនេះ វាច្បាស់ណាស់ថានៅក្នុងប្រភេទមួយមានភាពខុសប្លែកគ្នានៅក្នុងកម្រិតនៃសក្តានុពលបន្តពូជ។ ឧទាហរណ៍ នៅក្នុងលក្ខខណ្ឌនៃដំណាំចម្រុះ ទម្រង់ C មានសក្តានុពលបន្តពូជខ្ពស់បំផុត ខណៈដែលទម្រង់ A មានកម្រិតទាបបំផុត។
ជាចុងក្រោយ ទិន្នន័យពីតារាងទាំងពីរបង្ហាញថា ដំណាំក្រាស់មានអត្រាមរណៈច្រើនជាង ខណៈដំណាំក្រៀមមានតិចជាង។ ការមានកូនក៏ផ្លាស់ប្តូរតាមរបៀបដូចគ្នាដែរ។ ទិន្នន័យរបស់ Sukachev បង្ហាញថាសក្តានុពល biotic នៃទម្រង់ intraspecific មិនដូចគ្នាទេ ហើយជាលទ្ធផល ចំនួនប្រជាជននៃប្រភេទសត្វពិតជាមានក្រុមជីវសាស្រ្តមិនស្មើគ្នា។ សម្ភារៈដែលបានបង្ហាញក៏បង្ហាញផងដែរថានៅក្នុងប្រភេទសត្វមានការតស៊ូសម្រាប់អត្ថិភាពដែលបណ្តាលឱ្យមានការលុបបំបាត់ដោយជ្រើសរើស កំឡុងពេលដែលទម្រង់ដែលមានតិចបំផុត ក្រោមលក្ខខណ្ឌដែលបានផ្តល់ឱ្យ សក្តានុពលជីវសាស្រ្ត ពោលគឺ អ្នកដែលសម្របខ្លួនបានតិចតួចបំផុតត្រូវបានបំផ្លាញ។ ជាចុងក្រោយ ទិន្នន័យរបស់ Sukachev សង្កត់ធ្ងន់ថា ការរស់រានមានជីវិតរបស់អ្នកស័ក្តិសមបំផុត (អ្នកដែលមានសក្ដានុពលជីវសាស្ត្រខ្ពស់បំផុត) កើតឡើងមិនមែនតាមរយៈការជ្រើសរើសរបស់ពួកគេនោះទេ ប៉ុន្តែតាមរយៈការសម្លាប់ចោលនូវអ្វីដែលសមបំផុតនោះ។
Interspecies ប្រយុទ្ធសម្រាប់អត្ថិភាពក៏អាចមានភាពខ្លាំងក្លាផងដែរ។ ឧទាហរណ៍មួយចំនួននៃវាត្រូវបានផ្តល់ឱ្យខាងលើ។ ក្នុងករណីមួយចំនួន ពោលគឺប្រសិនបើផលប្រយោជន៍នៃប្រភេទសត្វមានភាពស្និទ្ធស្នាលទៅវិញទៅមក អាំងតង់ស៊ីតេនៃការតស៊ូអន្តរជាក់លាក់គឺមិនតិចជាងការតស៊ូក្នុងអន្តរជាក់លាក់នោះទេ។ ជាឧទាហរណ៍ ការប្រកួតប្រជែងខ្លាំងត្រូវបានគេសង្កេតឃើញរវាងសត្វក្រៀលពីរប្រភេទ - ម្រាមជើងតូចចង្អៀតខាងកើត (Astacus leptodactylus) និងម្រាមជើងធំទូលាយ (A. astacus) ជាមួយនឹងការផ្លាស់ទីលំនៅទីពីរ។
សូម្បីតែរវាងប្រភេទនៃក្រុមជាប្រព័ន្ធផ្សេងគ្នា ការប្រកួតប្រជែងគឺខ្ពស់ណាស់។ ឧទាហរណ៍ Zakarian (1930) បានសង្កេតឃើញថា រុក្ខជាតិ petrosimonia (P. brachiata) ជាក្បួនបានផ្លាស់ទីលំនៅប្រភេទសត្វផ្សេងទៀតដែលរីកលូតលាស់នៅក្នុងដីពិសោធន៍ដូចគ្នា។ ដូច្នេះនៅក្នុងការសង្កេតមួយនៅក្នុងខែមីនា Petrosimonia និងប្រភេទសត្វពីរទៀតបានកើនឡើងនៅក្នុងតំបន់ដូចគ្នា - Salsoda crassa និង Sueda splendens ។ ខាងក្រោមនេះត្រូវបានរាប់: 64 បុគ្គលនៃ Petrosiionia, 126 - S. crassa និង 21 - S. splendens ។ នៅរដូវស្លឹកឈើជ្រុះមានតែ Petrosimony ប៉ុណ្ណោះ។ ដូច្នេះនៅក្រោមលក្ខខណ្ឌនៃស្ថានីយ៍ដូចគ្នាការប្រកួតប្រជែងខ្លាំងកើតឡើងរវាងប្រភេទសត្វ។ មានតែនៅពេលដែលប្រភេទសត្វមានភាពខុសប្លែកគ្នាយ៉ាងខ្លាំងនៅក្នុងតម្រូវការរបស់ពួកគេ ទើបការប្រកួតប្រជែងរវាងពួកវាចុះខ្សោយ។ បន្ទាប់មកច្បាប់ (ដាវីន) នៃផលបូកដ៏ធំបំផុតនៃជីវិតជាមួយនឹងភាពចម្រុះដ៏អស្ចារ្យបំផុតចូលមកលេង។
វាគួរតែត្រូវបានចងចាំក្នុងចិត្តថា "ការតស៊ូអន្តរប្រភេទ" គឺមិនតែងតែចាំបាច់តិចជាង "ការតស៊ូអន្តរជាក់លាក់" នោះទេ។ អាំងតង់ស៊ីតេនៃការប្រកួតប្រជែងត្រូវបានកំណត់ដោយកត្តាជាច្រើន និងជាចម្បងដោយកម្រិតនៃភាពជិតនៃ niches ដែលកាន់កាប់។ ប្រសិនបើប្រភេទសត្វពីរកាន់កាប់កន្លែងដូចគ្នា នោះការប្រកួតប្រជែងរវាងពួកវានឹងមានលក្ខណៈធម្មជាតិនៃ "ការតស៊ូមិនជាក់លាក់"។ Gause (1935) បានសិក្សាករណីស្រដៀងគ្នាមួយ។ ciliates ពីរគឺ Paramaecium aurelia និង Glaucoma scintillans ត្រូវបានណែនាំទៅក្នុង "មីក្រូកូស" ។ ប្រសិនបើ P. aurelia ត្រូវបានលើកឡើងដោយឡែកពីគ្នា ចំនួនបុគ្គលកើនឡើងដល់កម្រិតឆ្អែតជាក់លាក់មួយ។ រឿងដដែលនេះកើតឡើងនៅក្នុងវប្បធម៌ដាច់ស្រយាលនៃជំងឺដក់ទឹកក្នុងភ្នែក។ ប្រសិនបើ ciliates ទាំងពីរចិញ្ចឹមនៅក្នុង microcosm នោះ ជំងឺដក់ទឹកក្នុងភ្នែកដែលមានអត្រាបន្តពូជខ្ពស់ គ្រប់គ្រងដើម្បីចាប់យកធនធានអាហារទាំងអស់នៅពេលដែល paramecium ទើបតែចាប់ផ្តើមលូតលាស់ជាលេខ ហើយជាលទ្ធផល ក្រោយមកទៀតត្រូវបានជំនួសទាំងស្រុង។ លទ្ធផលស្រដៀងគ្នានេះកើតឡើងនៅក្នុងវប្បធម៌ដែលមានពីរប្រភេទនៃ paramecia ហើយ P. aurelia បានផ្លាស់ប្តូរទាំងស្រុងនូវប្រភេទសត្វមួយផ្សេងទៀតដែលប្រើប្រាស់ធនធានអាហារមិនសូវមានផលិតភាព - P. caudatum ។ ទោះជាយ៉ាងណាក៏ដោយភាពស្មុគស្មាញមួយកើតឡើងនៅទីនេះដែលគុណសម្បត្តិនៃប្រភេទសត្វមួយលើប្រភេទមួយផ្សេងទៀតដូចដែលបានបញ្ជាក់ខាងលើ (សម្រាប់ទំនាក់ទំនងរវាងសត្វកណ្តុរ) អាស្រ័យលើលក្ខខណ្ឌបរិស្ថាន។ នៅក្នុងការពិសោធន៍របស់ Gause វាបានប្រែក្លាយថាប្រសិនបើ microcosm មានផលិតផលកាកសំណល់នៃ microorganisms រស់នៅក្នុងវានោះ P. aurelia ឈ្នះ។ ប្រសិនបើ microcosm ត្រូវបានលាងសម្អាតជាមួយនឹងដំណោះស្រាយអំបិលសុទ្ធនោះ P. caudatum អាចផ្លាស់ទីលំនៅ P. aurelia ។
ឥឡូវនេះ ចូរយើងបន្តទៅប្រភេទសត្វដែលមានលក្ខណៈពិសេសផ្សេងៗគ្នា។ paramecia ពីរត្រូវបានគេដាក់នៅក្នុង microcosm - P. aurelia និង P. bursaria ។ ប្រភេទទីពីរមានពណ៌ងងឹត អាស្រ័យលើសារាយ symbiotic ដែលរស់នៅក្នុងប្លាស្មារបស់វា។ សារាយបញ្ចេញអុកស៊ីហ៊្សែន (ក្នុងពន្លឺ) ហើយនេះធ្វើឱ្យ P. bursaria មិនសូវពឹងផ្អែកលើអុកស៊ីសែនបរិស្ថាន។ វាអាចមានដោយសេរីនៅផ្នែកខាងក្រោមនៃបំពង់សាកល្បង ដែលជាកន្លែងដែលកោសិកាមេដំបែប្រមូលផ្តុំ។ នេះគឺជាអ្វីដែល ciliates ចិញ្ចឹម។ P. aurelia គឺស្រលាញ់អុកស៊ីសែនច្រើនជាង (oxyphilic) ហើយស្នាក់នៅ ផ្នែកខាងលើបំពង់សាកល្បង ទម្រង់ទាំងពីរត្រូវបានប្រើប្រាស់ដោយផ្សិត និងបាក់តេរី ប៉ុន្តែទម្រង់មុនត្រូវបានប្រើយ៉ាងមានប្រសិទ្ធភាពជាងដោយ P. bursaria និងក្រោយមកដោយ P. aurelia ។ ដូច្នេះ niches របស់ពួកគេមិនស្របគ្នា។ តួលេខបង្ហាញថានៅក្រោមលក្ខខណ្ឌទាំងនេះ ការរួមរស់ជាអចិន្ត្រៃយ៍នៃប្រភេទទាំងពីរគឺអាចធ្វើទៅបាន។ ដូច្នេះ ដូចដែលយើងឃើញ ទិន្នន័យពិសោធន៍បញ្ជាក់ពីជំហររបស់ Darwin អំពីការថយចុះនៃអាំងតង់ស៊ីតេនៃការប្រកួតប្រជែងជាមួយនឹងភាពខុសគ្នានៃចំណាប់អារម្មណ៍ (ភាពខុសគ្នានៃតួអក្សរ) ហើយដោយហេតុនេះ អត្ថប្រយោជន៍នៃភាពខុសគ្នា។
ឧទាហរណ៍បុរាណនៃការតស៊ូដើម្បីអត្ថិភាពគឺជាទំនាក់ទំនងដែលកើតឡើងរវាងប្រភេទដើមឈើផ្សេងៗគ្នានៅក្នុងព្រៃ។ នៅក្នុងព្រៃ ការប្រកួតប្រជែងរវាងដើមឈើត្រូវបានគេសង្កេតឃើញយ៉ាងងាយស្រួល ក្នុងអំឡុងពេលដែលបុគ្គលមួយចំនួនយល់ថាខ្លួនឯងស្ថិតក្នុងទីតាំងដែលមានគុណសម្បត្តិ ចំណែកអ្នកផ្សេងទៀតស្ថិតក្នុងកម្រិតនៃការគៀបសង្កត់ផ្សេងៗគ្នា។

នៅក្នុងព្រៃឈើមាន៖ 1) ដើមលេចធ្លោ (I), 2) ដើមលេចធ្លោដែលមានមកុដមិនសូវលូតលាស់ល្អ (II), 3) ដើមលេចធ្លោដែលមានមកុដនៅដំណាក់កាលដំបូងនៃការ degeneration (III), 4) ដើមដែលត្រូវបានគេសង្កត់ (IV), 5) ប្រចៀវស្លាប់និងស្លាប់ (V). ប្រភេទដើមឈើខុសៗគ្នាក្នុងជីវភាពរស់នៅខុសគ្នាយ៉ាងច្បាស់ផ្លាស់ប្តូរគ្នាទៅវិញទៅមក ដូច្នេះ នៅប្រទេសដាណឺម៉ាក ការផ្លាស់ទីលំនៅរបស់ដើមប៊ីចដោយដើមប៊ីចត្រូវបានតាមដាន។ ព្រៃ birch សុទ្ធត្រូវបានអភិរក្សតែនៅតំបន់វាលខ្សាច់ និងដីខ្សាច់ប៉ុណ្ណោះ ប៉ុន្តែកន្លែងដែលដីមានលក្ខណៈសមរម្យសម្រាប់ដើមប៊ីច វាបានច្រានចេញនូវដើមប៊ីច។ នៅក្នុងស្ថានភាពនេះ វាអាចរស់នៅបានយូរអង្វែង ប៉ុន្តែនៅទីបំផុតស្លាប់ ដោយសារដើមប៊ីចមានអាយុវែងជាងវា ហើយមកុដរបស់វាមានថាមពលខ្លាំងជាង។ លើសពីនេះទៀតដើមប៊ីចលូតលាស់នៅក្រោមដំបូលនៃ birch ខណៈពេលដែលក្រោយមកទៀតមិនអាចដុះនៅក្រោមដំបូលនៃដើមប៊ីច។
ការជ្រើសរើសធម្មជាតិ
ពីការតស៊ូដើម្បីអត្ថិភាព ការជ្រើសរើសធម្មជាតិកើតឡើងជាលទ្ធផល។ ដាវីនមិនមានឱកាសពឹងផ្អែកលើការសង្កេតដោយផ្ទាល់ដែលបញ្ជាក់ដោយផ្ទាល់នូវសកម្មភាពនៃការជ្រើសរើសធម្មជាតិនោះទេ។ ដើម្បីបង្ហាញវា គាត់បានប្រើឧទាហរណ៍ "ការស្រមើលស្រមៃ" ដូចដែលគាត់ផ្ទាល់បានបង្ហាញ។ ពិតហើយ ឧទាហរណ៍ទាំងនេះដកដង្ហើមជីវិតខ្លួនឯង។ ទោះជាយ៉ាងណាក៏ដោយ ពួកគេមិនមែនជាភស្តុតាងច្បាស់លាស់នៃការជ្រើសរើសធម្មជាតិនោះទេ។ ក្រោយមក ស្ថានការណ៍បានផ្លាស់ប្តូរ ហើយការងារបន្តិចម្តងៗចាប់ផ្តើមលេចឡើង ដែលការពិតនៃការជ្រើសរើសធម្មជាតិត្រូវបានបញ្ជាក់។
ការពិតដែលគាំទ្រទ្រឹស្តីនៃការជ្រើសរើសធម្មជាតិអាចបែងចែកជាពីរក្រុម៖ ភស្តុតាងដោយប្រយោលនៃការជ្រើសរើសធម្មជាតិ និងភស្តុតាងផ្ទាល់។
ភស្តុតាងប្រយោលនៃការជ្រើសរើសធម្មជាតិ. នេះរួមបញ្ចូលទាំងក្រុមនៃអង្គហេតុដែលទទួលបានការពេញចិត្តបំផុតរបស់ពួកគេ ឬសូម្បីតែការពន្យល់តែមួយគត់នៅលើមូលដ្ឋាននៃទ្រឹស្តីនៃការជ្រើសរើសធម្មជាតិ។ ពីការពិតស្រដៀងគ្នាមួយចំនួនធំ យើងនឹងផ្តោតលើចំណុចខាងក្រោម៖ ពណ៌ការពារ និងរូបរាង និងបាតុភូតនៃការធ្វើត្រាប់តាម លក្ខណៈពិសេសនៃការសម្របខ្លួននៃផ្កា entomophilous, ornithophilous និង theriophilous, តួអក្សរសម្របខ្លួននៃសត្វល្អិតកោះ, អាកប្បកិរិយាសម្របខ្លួន, របៀប! ភស្តុតាងនៃការជ្រើសរើស។
1. គាំទ្រពណ៌និងរូបរាង. តាមរយៈការដាក់ពណ៌ និងរាងការពារ ឬពណ៌គ្រីប និងរូបរាង យើងមានន័យថាភាពស្រដៀងគ្នានៃសារពាង្គកាយ (ពណ៌ ឬរាង) ជាមួយនឹងវត្ថុក្នុងបរិយាកាសរស់នៅធម្មតារបស់វា។
បាតុភូតនៃភាពស្រដៀងគ្នានៃគ្រីបគឺរីករាលដាលនៅក្នុងធម្មជាតិ។ សូមក្រឡេកមើលឧទាហរណ៍មួយចំនួននៃការដាក់ពណ៌ និងរូបរាងគ្រីប។
អ្នកសត្វវិទ្យាជនជាតិរុស្សី V.A. Wagner (1901) បានពិពណ៌នាអំពីសត្វពីងពាង (Drassus polihovi) ដែលស្ថិតនៅលើមែកឈើ ហើយមានលក្ខណៈស្រដៀងនឹងដើមមែក។ ពោះរបស់វាត្រូវបានគ្របដណ្ដប់ដោយផ្នត់ដែលស្រដៀងទៅនឹងជញ្ជីងតំរងនោម។ សត្វពីងពាងធ្វើចលនាខ្លី និងលឿន ដោយសន្មត់ថាសម្រាកភ្លាមៗ និងធ្វើត្រាប់តាមក្រលៀន។ ដូច្នេះភាពស្រដៀងគ្នានៃគ្រីបត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងអាកប្បកិរិយាគ្រីប (ឥរិយាបថសម្រាក) - ការពិតដែលជាលក្ខណៈមិនធម្មតានៃបាតុភូតដែលបានពិពណ៌នាដែលរីករាលដាលក្នុងចំណោមសត្វរួមទាំងសត្វឆ្អឹងខ្នង។ ដូច្នេះ សត្វស្លាបព្រៃជាច្រើនមានពណ៌ និងលម្អដើម្បីផ្គូផ្គងពណ៌ និងផ្ទៃនៃសំបកឈើ។ សត្វស្លាបបែបនេះ (ឧទាហរណ៍ សត្វទីទុយ សត្វទីទុយ ឥន្ទ្រី សត្វទីទុយ ក្អែករាត្រី ភីកា ជាដើម) គឺមើលមិនឃើញទាំងស្រុងនៅក្នុងទីតាំងសម្រាក។ នេះអនុវត្តជាពិសេសចំពោះស្ត្រី។ ភាពស្រដៀងនឹងសំបកឈើរបស់ពួកវាគឺមានសារៈសំខាន់ខ្លាំងសម្រាប់ហេតុផលដែលថាវាជាធម្មតាជាស្ត្រីដែលអង្គុយនៅលើពង ឬការពារកូនមាន់។ ដូច្នេះក្នុងករណីដែលបុរស ប្រភេទព្រៃឈើ(ឧទាហរណ៍ ក្ងោកខ្មៅ និង capercaillie) មានភាពខុសប្លែកគ្នាយ៉ាងល្អពីគ្នាទៅវិញទៅមក ពណ៌ស្រីរបស់ពួកគេមានពណ៌ស្រដៀងគ្នាខ្លាំងណាស់ (ឯកសណ្ឋាន)។ សម្រាប់ហេតុផលដូចគ្នា ជាឧទាហរណ៍ នៅក្នុងសត្វស្លាបទូទៅ (Phasianus colchicus) ពូជភូមិសាស្រ្តពណ៌គឺជាលក្ខណៈរបស់បុរសតែប៉ុណ្ណោះ ខណៈពេលដែលញីនៃប្រភេទរងភូមិសាស្ត្រទាំងអស់នៃបក្សីនេះមានពណ៌ស្រដៀងនឹងការការពារ។ បាតុភូតស្រដៀងគ្នានេះត្រូវបានគេសង្កេតឃើញនៅក្នុងសត្វដទៃទៀត។
លំនាំនៃពណ៌គ្រីប. លក្ខណៈពិសេសចម្បងនៃបាតុភូតគ្រីបគឺថាផ្នែកទាំងនោះនៃរាងកាយដែលត្រូវបានប៉ះពាល់នឹងភ្នែករបស់សត្វមំសាសីមានពណ៌សម្ងាត់។ ដូច្នេះ ជាឧទាហរណ៍ នៅក្នុងមេអំបៅដែលបត់ស្លាបរបស់ពួកគេក្នុងលក្ខណៈដូចដំបូល (ជាលទ្ធផលដែលផ្នែកខាងលើនៃស្លាបខាងមុខប្រឈមមុខនឹងអ្នកសង្កេតការណ៍) ពណ៌គ្រីបគឺតែងតែមានវត្តមានយ៉ាងជាក់លាក់នៅផ្នែកខាងលើនេះ។ ផ្នែកដែលនៅសល់នៃស្លាបដែលគ្របដណ្ដប់ (នៅក្នុងទីតាំងសម្រាក) ហើយមើលមិនឃើញ អាចនិងជាញឹកញាប់មានពណ៌ភ្លឺ។ ជាឧទាហរណ៍ សត្វប្រចៀវដែលមានស្លាបក្រហម (Catoeala nupta និងប្រភេទសត្វដទៃទៀត) មានឆ្នូតពណ៌ក្រហមភ្លឺនៅលើស្លាបខាងក្រោយរបស់វា។ ក្នុងអំឡុងពេលហោះហើរយ៉ាងលឿននៃមេអំបៅនេះ ពួកវាបញ្ចេញពន្លឺនៅចំពោះមុខអ្នក។ ទោះជាយ៉ាងណាក៏ដោយ នៅពេលដែលវាអង្គុយនៅលើសំបកឈើ នោះពណ៌គ្រីប (ដើម្បីផ្គូផ្គងពណ៌សំបកឈើ) ស្លាបខាងមុខត្រួតលើស្លាបខាងក្រោយភ្លឺដូចជាដំបូល ហើយមេអំបៅក៏បាត់ពីទិដ្ឋភាព លុះត្រាតែខ្សែកោងដែលខូចនៃការហោះហើររបស់វាត្រូវបាត់បង់។ ការមើលឃើញ។ បាតុភូតនេះកាន់តែមានប្រសិទ្ធភាពនៅក្នុង Kallima ដែលក្នុងនោះភាពស្រដៀងគ្នានៃគ្រីបឈានដល់ឯកទេសខ្ពស់។
ដូចមេអំបៅពេលថ្ងៃទាំងអស់ ស្លាបរបស់ពួកវាបត់នៅខាងក្រោយខ្នង មិនមែនដូចមេអំបៅទេ (ដូចមេអំបៅ) ប៉ុន្តែស្របគ្នាទៅវិញទៅមក។ ដូច្នេះហើយ នៅក្នុងការសម្រាកមួយ ផ្នែកខាងលើនៃស្លាបត្រូវបានលាក់ ហើយផ្នែកខាងក្រោមត្រូវប្រឈមមុខនឹងអ្នកសង្កេត។ ក្នុងករណីនេះ ផ្នែកខាងលើដែលលាក់មានពណ៌ភ្លឺដែលអាចមើលឃើញក្នុងអំឡុងពេលហោះហើរ (ឧទាហរណ៍ ឆ្នូតពណ៌លឿងនៅលើផ្ទៃខាងក្រោយពណ៌ខៀវ) ហើយផ្នែកខាងក្រោមខាងក្រៅមានពណ៌សំខាន់។ លោក Wallace ដែលបានសង្កេតមើល callim នៅលើកោះ។ ស៊ូម៉ាត្រា បង្ហាញថា វាគ្រប់គ្រាន់សម្រាប់មេអំបៅអង្គុយលើមែកឈើ ហើយវាបានបាត់បង់ ដែលត្រូវបានសម្របសម្រួលមិនត្រឹមតែដោយពណ៌សម្ងាត់នៃស្លាបប៉ុណ្ណោះទេ ប៉ុន្តែក៏មានលំនាំ និងរូបរាងគ្រីបរបស់វា ខុសពីស្លឹកឈើដែរ។ ជាមួយ petiole មួយ។
ដូច្នេះ ការលាបពណ៌គ្រីប ជាដំបូង មានវត្តមាននៅក្នុងបុគ្គលទាំងនោះ ដែលវាមានប្រយោជន៍ជាពិសេស (ឧទាហរណ៍ មនុស្សស្រី) ហើយទីពីរ វាវិវត្តន៍នៅក្នុងផ្នែកទាំងនោះនៃរាងកាយ ដែលប៉ះពាល់នឹងភ្នែករបស់សត្វមំសាសី (កន្លែងដែលវាត្រូវការ។ ជាមធ្យោបាយក្លែងបន្លំ)។ ទីបី បាតុភូតគ្រីបតែងតែត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងឥរិយាបថសម្រាក ពោលគឺជាមួយនឹងអាកប្បកិរិយាសំខាន់ដែលបង្កើនប្រសិទ្ធភាពគ្រីបនៃការបិទបាំង (Oudemans, 1903)។
ទោះជាយ៉ាងណាក៏ដោយ បាតុភូតដ៏គួរឱ្យកត់សម្គាល់ទាំងនេះមិនបញ្ចប់នៅទីនោះទេ។ សត្វល្អិតដំបង (Phasmidae) ដែលត្រូវបានសិក្សាដំបូងដោយ Bates (1862) ត្រូវបានគេស្គាល់ថាមានរូបរាងស្រដៀងទៅនឹង knotweeds ។ ឥរិយាបថសម្រាក (អាកប្បកិរិយារិះគន់) បន្ថែមភាពស្រដៀងគ្នានេះ។ ប្រសិនបើអ្នកប៉ះដំបងឈើ វាយោលមួយរយៈ ដូចជាស្មៅដែលបក់ដោយខ្យល់ (ចលនាការពារ)។ ប្រសិនបើអ្នករើសសត្វល្អិតនៅក្នុងដៃរបស់អ្នក វានឹងធ្លាក់ចូលទៅក្នុងស្ថានភាពនៃ thanatosis (ការឆ្លុះបញ្ជាំងបណ្ដោះអាសន្ន និងងាយស្រួលបញ្ចប់ភាពអសកម្ម)។ ក្នុងករណីនេះ សត្វល្អិតដំបងបត់ជើងនៅតាមដងខ្លួនរបស់វា ហើយក្លាយទៅជាមិនអាចសម្គាល់បានទាំងស្រុងពីស្មៅស្ងួត។ បាតុភូត Thanatosis គឺជាលក្ខណៈនៃសត្វល្អិតជាច្រើន។
2. ការធ្វើត្រាប់តាម. នេះគឺជាឈ្មោះដែលផ្តល់ភាពស្រដៀងគ្នានៃសត្វមួយចំនួន (អ្នកធ្វើត្រាប់តាម ឬអ្នកត្រាប់តាម) ជាមួយសត្វផ្សេងទៀតដែលមានអត្ថន័យនៃ "គំរូ" ហើយ "អ្នកត្រាប់តាម" ទទួលបានអត្ថប្រយោជន៍មួយឬផ្សេងទៀតពីភាពស្រដៀងគ្នាជាមួយ "គំរូ" ។ ការធ្វើត្រាប់តាមគឺរីករាលដាលក្នុងចំណោមសត្វល្អិត ជាពិសេសនៅក្នុងធម្មជាតិរុស្ស៊ីរបស់យើង។ សត្វរុយខ្លះមកពីគ្រួសារ Syrphidae ធ្វើត្រាប់តាមសត្វស្វា និងសត្វឃ្មុំ ខណៈដែលសត្វល្អិតជាច្រើនដែលជាកម្មសិទ្ធិរបស់ការបញ្ជាទិញផ្សេងៗ ក៏ដូចជាសត្វពីងពាងខ្លះមានទំនាក់ទំនងជីវសាស្រ្តជាមួយស្រមោច និងបង្កើតជាក្រុមដែលគេហៅថា myrmecophilesមានលក្ខណៈស្រដៀងនឹងស្រមោច។ មេអំបៅខ្លះធ្វើត្រាប់តាមអ្នកដទៃ ជាសត្វដែលមិនអាចបរិភោគបាន ដែលពួកវាហើរជាមួយគ្នា។
មេអំបៅ Papilio dardanus ត្រូវបានរកឃើញនៅក្នុងទ្វីបអាហ្រ្វិក ដែលមានជួរធំណាស់ ចាប់ពី Abyssinia រហូតដល់ Cape Colony រួមបញ្ចូល និងពីច្រាំងភាគខាងកើត រហូតដល់សេណេហ្គាល់ និងឆ្នេរមាស។ លើសពីនេះទៀត P. dardanus ត្រូវបានរកឃើញនៅក្នុងប្រទេសម៉ាដាហ្គាស្ការ។ ទម្រង់ដែលរស់នៅលើកោះនេះជាទូទៅមានលក្ខណៈធម្មតាសម្រាប់ genus នៅក្នុងលំនាំ និងវណ្ឌវង្កនៃស្លាប ដែលនឹកឃើញដល់ swallowtails រុស្ស៊ីរបស់យើង។
រូបភាពខុសគ្នាទាំងស្រុងត្រូវបានគេសង្កេតឃើញនៅលើទ្វីបអាហ្រ្វិក។ នៅទីនេះ លើកលែងតែ Abyssinia ដែលជាកន្លែងញីធម្មតារបស់ P. dardanus ត្រូវបានរកឃើញ ភាពចម្រុះនៃប្រភេទសត្វនៅក្នុងសំណួរត្រូវបានអង្កេតឃើញ។ polymorphism នេះត្រូវបានភ្ជាប់ក្នុងករណីនេះជាមួយការធ្វើត្រាប់តាម។
នៅអាហ្រ្វិកខាងត្បូងគឺនៅក្នុង Cape Colony ស្ត្រីនៃ P. dardanus ត្រូវបានកែប្រែទាំងស្រុង។ ស្លាបរបស់ពួកគេខ្វះតុល្យភាព និងបញ្ឆោតស្រដៀងនឹងស្លាបរបស់មេអំបៅក្នុងស្រុកមួយទៀតគឺ Amauris echeria (ក៏គ្មានតុល្យភាពផងដែរ)៖
នេះគឺជា "គំរូ" ដែលជនជាតិដើម P. dardanus យកតម្រាប់តាម។ លើសពីនេះទៅទៀត A. echeria ក៏រស់នៅក្នុង Natal ហើយបង្កើតទម្រង់ក្នុងស្រុកពិសេសមួយនៅទីនេះ ដែលត្រូវបានតភ្ជាប់ដោយការផ្លាស់ប្តូរមួយចំនួនជាមួយនឹងទម្រង់ Cape នៃប្រភេទដូចគ្នា។ ហើយឥឡូវនេះ ញីរបស់ P. dardanus ធ្វើត្រាប់តាមប្រភេទសត្វនេះ ផ្តល់នូវទម្រង់អន្តរកាលស្របគ្នា (ពី Cape ទៅ Natal) ដោយយកតម្រាប់តាមទម្រង់អន្តរកាលនៃ "គំរូ" ។
ទោះជាយ៉ាងណាក៏ដោយបាតុភូតដែលបានពិពណ៌នាមិនត្រូវបានកំណត់ចំពោះរឿងនេះទេ។ បន្ថែមពីលើ A. echeria មេអំបៅពីរក្បាលទៀតហើរនៅតំបន់ Cape Colony: A. niavius និង Danais chrysippus ។ ដូច្នោះហើយ ស្ត្រីក្នុងស្រុកនៃ P. dardanus បង្កើតឱ្យមានទម្រង់ត្រាប់តាមពីរបន្ថែមទៀត។ មួយក្នុងចំណោមពួកគេធ្វើត្រាប់តាម D. chrysippus និងមួយទៀត A. niavius ។
ដូច្នេះ P. dardanus មានទម្រង់ស្រីជាច្រើនដែលធ្វើត្រាប់តាម "គំរូ" ជាច្រើនគឺទម្រង់ Cape និង Natal នៃ A. echeria ។ A. niavius, Danais chrysippus ។
សំណួរធម្មជាតិកើតឡើង៖ តើអ្វីជាអត្ថន័យជីវសាស្រ្តនៃការធ្វើត្រាប់តាមទាំងនេះ? វាត្រូវបានគេរកឃើញថា "គំរូ" ជាកម្មសិទ្ធិរបស់មេអំបៅដែលមិនអាចបរិភោគបាន។ សត្វល្អិតជៀសវាងពួកវាក្នុងករណីណាក៏ដោយ។ ក្នុងពេលជាមួយគ្នានេះ សត្វស្លាបពិតជាត្រូវបានតម្រង់ទិសដោយចក្ខុវិស័យ ហើយពណ៌ជាក់លាក់មួយ (និងរូបរាង) នៃស្លាបមេអំបៅត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការឆ្លុះបញ្ចាំងតាមលក្ខខណ្ឌជាមួយនឹងអារម្មណ៍មិនល្អសម្រាប់សត្វស្លាប (ជាក់ស្តែង រសជាតិ)។ ជាលទ្ធផល "អ្នកត្រាប់តាម" (ក្នុងករណីនេះស្ត្រីនៃ P. dardanus) ខណៈពេលដែលនៅសល់នៅក្នុងការពិតអាចបរិភោគបានប៉ុន្តែក្នុងពេលតែមួយមានភាពស្រដៀងគ្នាជាមួយ "គំរូ" ដែលមិនអាចបរិភោគបានគឺក្នុងកម្រិតជាក់លាក់មួយដែលត្រូវបានការពារពីការវាយប្រហារដោយសត្វស្លាបដែល " ខុស” ពួកគេសម្រាប់ក្រោយ។
3. ការពន្យល់អំពីបាតុភូតគ្រីប និងការធ្វើត្រាប់តាម ដោយផ្អែកលើទ្រឹស្តីនៃការជ្រើសរើសធម្មជាតិ. បាតុភូតនៃទម្រង់សម្ងាត់ និងឥរិយាបទ ក៏ដូចជាបាតុភូតនៃការធ្វើត្រាប់តាមដែលបានពិពណ៌នាខាងលើ គឺរីករាលដាលយ៉ាងខ្លាំងនៅក្នុងក្រុមផ្សេងៗនៃសារពាង្គកាយ ដែលមនុស្សម្នាក់មិនអាចជួយបាន ប៉ុន្តែមើលឃើញនៅក្នុងគំរូជាក់លាក់មួយដែលត្រូវការការពន្យល់ពីមូលហេតុ។ ក្រោយមកទៀតត្រូវបានសម្រេចទាំងស្រុងលើមូលដ្ឋាននៃទ្រឹស្តីនៃការជ្រើសរើសធម្មជាតិ។ ទោះយ៉ាងណាក៏ដោយ ការពន្យល់ផ្សេងទៀតត្រូវបានស្នើឡើង។ អ្នកស្រាវជ្រាវខ្លះទទួលស្គាល់ថា ជាឧទាហរណ៍ ការលាបពណ៌ លំនាំ និងរូបរាងជាគ្រីប គឺជាលទ្ធផលនៃឥទ្ធិពលនៃកត្តារូបវិទ្យា-គីមី ការធ្វើលំហាត់ប្រាណ ឬផលវិបាកនៃកត្តាផ្លូវចិត្តពិសេស។ល។
ចូរយើងពិចារណាការសន្មត់ទាំងនេះ។ ជាឧទាហរណ៍ តើវាអាចទៅរួចទេក្នុងការសន្មត់ថា "បុព្វបុរស" callim "អនុវត្ត" ក្នុងភាពស្រដៀងនឹងស្លឹកឈើ ឬស្រីរបស់ P. dardanus ក្នុងភាពស្រដៀងនឹង "គំរូ" ដែលត្រូវគ្នា? ភាពមិនសមហេតុផលនៃ "ការពន្យល់" បែបនេះគឺបង្ហាញឱ្យឃើញដោយខ្លួនឯង។ វាមិនសមហេតុផលទេក្នុងការសន្មត់ថាសំណួរគឺអំពីឥទ្ធិពលនៃអាកាសធាតុ សីតុណ្ហភាព សំណើម អាហារ។ល។
តើកត្តាទាំងនេះធ្វើឱ្យសត្វល្អិតដំបងមានលក្ខណៈស្រដៀងនឹងមែកឈើដោយរបៀបណា ហើយផ្កាខាត់ណាស្រដៀងនឹងស្លឹក? ហេតុអ្វីបានជាកត្តាទាំងនេះមានឥទ្ធិពលគ្រីបនៅលើផ្នែកខាងក្រោមនៃស្លាបរបស់ callima និងនៅផ្នែកខាងលើនៃស្លាបរបស់ខ្សែបូក្រហម? វាច្បាស់ណាស់ថាការប៉ុនប៉ងកាត់បន្ថយពណ៌ការពារ និងរូបរាង ឬការធ្វើត្រាប់តាមទៅនឹងឥទ្ធិពលសរីរវិទ្យាសុទ្ធសាធនៃកត្តាខាងក្រៅគឺគ្មានផលផ្លែទេ។ អ្នកគួរតែគិតអំពីការពិតដែលថាសត្វរុយនិងខ្សែបូមានពណ៌ការពារតែនៅលើផ្នែកទាំងនោះនៃស្លាបដែលប្រឈមមុខ (ក្នុងស្ថានភាពសម្រាក) ទៅនឹងបរិយាកាសខាងក្រៅ។ ផ្នែកដូចគ្នានៃស្លាបដែលត្រូវបានលាក់នៅក្នុងការសម្រាក, មិនត្រឹមតែមិនមាន ការឧបត្ថម្ភពណ៌ប៉ុន្តែផ្ទុយទៅវិញ ពួកគេមានលំនាំភ្លឺដែលទាក់ទាញភ្នែក។ នៅក្នុងមេអំបៅ crepuscular និងពេលយប់ ផ្នែកតូចមួយនៃស្លាបខាងក្រោយនៅតែអាចមើលឃើញនៅក្នុងទីតាំងសម្រាក។ ដូច្នេះហើយ វាគឺជាផ្នែកនៃស្លាបខាងក្រោយដែលមានពណ៌គ្រីប ខណៈពេលដែលនៅសល់នៃពួកវា ដែលលាក់ពីការក្រឡេកមើលសត្វស្លាបគ្មានសត្វល្អិត មិនមានពណ៌គ្រីបនេះទេ។
វាច្បាស់ណាស់ថានៅក្នុងករណីបែបនេះវាគ្រាន់តែជាការមិនសមហេតុផលក្នុងការនិយាយអំពីការធ្វើលំហាត់ប្រាណ ឥទ្ធិពលនៃអាហារ ពន្លឺ សីតុណ្ហភាព សំណើម។ល។ ដូចក្នុងឧទាហរណ៍ពីមុន។
ដូច្នេះ ប្រសិនបើបាតុភូតនៃភាពស្រដៀងគ្នា និងការធ្វើត្រាប់តាមសម្ងាត់គឺមិនអាចពន្យល់បានពីចំណុចនៃទិដ្ឋភាពដែលបានចង្អុលបង្ហាញនោះ ផ្ទុយទៅវិញ ពួកគេទទួលបានការពន្យល់ដែលពេញចិត្តនៅក្នុងពន្លឺនៃទ្រឹស្តីនៃការជ្រើសរើស។
តាមការពិត ពីកត្តាដែលបានពិពណ៌នាខាងលើ វាច្បាស់ណាស់ថាភាពស្រដៀងគ្នា និងការធ្វើត្រាប់តាមគ្រីបមានប្រយោជន៍សម្រាប់ម្ចាស់របស់វា។ រាល់ការផ្លាស់ប្តូរតំណពូជទាំងនោះដែលបណ្តាលឱ្យមានការលេចចេញនូវភាពស្រដៀងគ្នានៃកូដសម្ងាត់ត្រូវបានរក្សាទុកដោយសារតែអត្ថប្រយោជន៍របស់វា។ អាស្រ័យហេតុនេះ ពីមួយជំនាន់ទៅមួយជំនាន់ មានជម្រើសធម្មជាតិសម្រាប់គុណភាពសម្ងាត់។
ការធ្វើត្រាប់តាមត្រូវបានពន្យល់តាមរបៀបស្រដៀងគ្នា។ ជាឧទាហរណ៍ វាត្រូវបានគេរកឃើញថាស្ត្រីនៃបីប្រភេទដែលបានចង្អុលបង្ហាញខាងលើអាចផុសចេញពីពងស្វាសដែលមានទម្រង់ដូចគ្នានៃ P. dardanus ។ អាស្រ័យហេតុនេះ នៅក្នុងតំបន់ដែលបានផ្តល់ឱ្យ ទម្រង់ផ្សេងៗគ្នានៃស្ត្រី P. dardanus អាចលេចឡើង ប៉ុន្តែតាមពិត អ្នកដែលយកតម្រាប់តាមគំរូក្នុងស្រុកបានល្អជាងអ្នកផ្សេងទៀតនឹងត្រូវបានរក្សាទុក។ នៅសល់ បើទោះបីជាពួកគេបានបង្ហាញខ្លួនក៏ដោយ មានឱកាសរស់រានមានជីវិតទាបជាងច្រើន ដោយសារមិនមានគំរូដែលមិនអាចបរិភោគបាននៅក្នុងតំបន់ដែលបានផ្តល់ឱ្យ ដូច្នេះហើយ សត្វស្លាបនឹងបំផ្លាញអ្នកត្រាប់តាម “គ្មានមូលដ្ឋាន” បែបនេះ។
ការពន្យល់ទូទៅនេះតម្រូវឱ្យមានការឌិកូដមួយចំនួន។ ជាឧទាហរណ៍ ប្រសិនបើយើងព្យាយាមវិភាគភាពស្រដៀងគ្នានៃអក្សរកាត់នៃ callima ទៅនឹងស្លឹកមួយ នោះយើងនឹងរកឃើញភ្លាមៗថាវាត្រូវបានផ្សំឡើងដោយធាតុមួយចំនួនធំ។ ភាពស្រដៀងគ្នានៃ callima ជាមួយស្លឹកស្លឹកគឺលម្អិតមិនមែនទូទៅទេ។ នេះគឺជាទម្រង់ស្លឹកទូទៅនៃស្លាបបត់ លំនឹង ដែលនៅពេលបត់ត្រូវគ្នានឹងដើមស្លឹក បន្ទាត់មធ្យមនៃលំនាំគ្រីបនៃស្លាប ធ្វើត្រាប់តាមផ្នែកកណ្តាលនៃស្លឹក។ ធាតុនៃ venation នៅពេលក្រោយ; ចំណុចនៅលើស្លាប ការធ្វើត្រាប់តាមចំណុចផ្សិតនៅលើស្លឹក ការលាបពណ៌ទូទៅនៃផ្នែកខាងក្រោមនៃស្លាប ធ្វើត្រាប់តាមពណ៌នៃស្លឹកកាហ្វេស្ងួត ហើយចុងក្រោយគឺឥរិយាបថរបស់ callima ដោយប្រើភាពស្រដៀងនឹងស្លឹកគ្រីបរបស់វាជាមួយនឹងជំនួយ។ នៃឥរិយាបថសម្រាកសមរម្យ។
ធាតុទាំងអស់នៃការដាក់ពណ៌ ទម្រង់ និងអាកប្បកិរិយាមិនអាចកើតឡើងភ្លាមៗនោះទេ។ ដូចគ្នាដែរចំពោះករណីដែលបានពិពណ៌នាអំពីការធ្វើត្រាប់តាម។ ការបង្កើតភ្លាមៗនៃធាតុទាំងអស់នេះនឹងជាអព្ភូតហេតុមួយ។ ទោះជាយ៉ាងណាក៏ដោយ អព្ភូតហេតុមិនកើតឡើងទេ ហើយវាច្បាស់ណាស់ថា ធាតុគ្រីបរបស់ កាលីម៉ា ត្រូវបានផ្សំឡើងជាប្រវត្តិសាស្ត្រ។ តាមទស្សនៈនៃទ្រឹស្ដីនៃការជ្រើសរើស ភាពស្រដៀងគ្នានៃគ្រីប និងការធ្វើត្រាប់តាមបានកើតឡើងជាការចៃដន្យ ហើយលើសពីនេះទៅទៀត ភាពស្រដៀងគ្នាប្រហាក់ប្រហែល។ ទោះយ៉ាងណាក៏ដោយនៅពេលដែលវាកើតឡើងបន្ទាប់មកវាត្រូវបានបម្រុងទុកជាប្រយោជន៍។ ដោយបានបន្តឆ្លងកាត់ជំនាន់ជាច្រើន ភាពស្រដៀងគ្នានៃគ្រីបដំបូងត្រូវបានបង្ហាញនៅក្នុងបុគ្គលផ្សេងៗគ្នា ទៅតាមកម្រិតផ្សេងៗគ្នា និងក្នុងចំនួនធាតុផ្សេងៗគ្នា។ នៅក្នុងបុគ្គលមួយចំនួន ឥទ្ធិពលគ្រីបនៃលក្ខណៈដែលបានផ្តល់ឱ្យ (ឧទាហរណ៍ ពណ៌ស្លាប) ឬឥទ្ធិពលនៃភាពស្រដៀងគ្នាទៅនឹងទម្រង់ដែលមិនអាចបរិភោគបានគឺពេញលេញជាងអ្នកដទៃ។ ទោះជាយ៉ាងណា វាជារឿងធម្មជាតិប្រសិនបើមានមនុស្សប្រុងប្រយ័ត្ននៅក្នុងតំបន់នោះ។ បក្សីសត្វល្អិតបុគ្គលដែលមានឥទ្ធិពលខ្ពស់បំផុត និងច្រើនបំផុត មួយចំនួនធំសញ្ញាសម្ងាត់ ឬសញ្ញានៃការធ្វើត្រាប់តាម។
ដូច្នេះ ក្នុងជំនាន់ដ៏យូរមកហើយ ទម្រង់ដែលមានលក្ខណៈសម្ងាត់បំផុតបានរួចជីវិត។ ដូច្នេះតាមធម្មជាតិ ភាពស្រដៀងនឹងគ្រីប និងការធ្វើត្រាប់តាមត្រូវបានកែលម្អចាំបាច់។ សញ្ញាគ្រីបនីមួយៗត្រូវបានពង្រឹង ហើយចំនួននៃសញ្ញាសម្ងាត់បែបនេះបានប្រមូលផ្តុំ។ នេះជារបៀបដែលភាពស្មុគស្មាញនៃលក្ខណៈសម្ងាត់នៃ callima ដែលបានពិពណ៌នាខាងលើត្រូវបានបង្កើតឡើងជាប្រវត្តិសាស្ត្រ។ វាមិនធ្វើតាមអ្វីដែលបាននិយាយទេ ពិតណាស់ថាសារពាង្គកាយទាំងមូលគឺជាលទ្ធផលសាមញ្ញនៃការបូកសរុបនៃលក្ខណៈ។ ឥទ្ធិពលគ្រីបទិកពិតជាកកកុញ ប៉ុន្តែការប្រមូលផ្តុំនេះតែងតែត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការរីកធំធាត់ទូទៅនៃសារពាង្គកាយដែលជាលទ្ធផលនៃការរួមបញ្ចូលគ្នាតាមរយៈការឆ្លងកាត់។ បញ្ហានេះត្រូវបានពិភាក្សាដូចខាងក្រោម។
ទោះជាយ៉ាងណាក៏ដោយ ប្រសិនបើភាពស្រដៀងនឹងគ្រីប និងការធ្វើត្រាប់តាមត្រូវបានធ្វើឱ្យល្អឥតខ្ចោះនៅក្នុងដំណើរនៃប្រវត្តិសាស្រ្ត នោះយើងគួររំពឹងថាប្រភេទសត្វផ្សេងៗគ្នា ឧទាហរណ៍ ប្រភេទសត្វមេអំបៅ ក៏គួរតែស្ថិតនៅក្នុងយុគសម័យភូគព្ភសាស្ត្រនៅដំណាក់កាលផ្សេងៗគ្នានៃភាពឥតខ្ចោះនៃការសម្របខ្លួននេះ។ អ្វីដែលត្រូវបានរំពឹងទុកតាមទ្រឹស្តីគឺពិតជាត្រូវបានសង្កេតឃើញនៅក្នុងធម្មជាតិ។ តាមពិត ពណ៌ និងរូបរាងសំខាន់ៗត្រូវបានបង្ហាញជាមួយនឹងកម្រិតនៃភាពល្អឥតខ្ចោះខុសៗគ្នានៅក្នុងប្រភេទផ្សេងៗគ្នា។ ក្នុងករណីខ្លះសត្វល្អិតមិនមានតួអក្សរគ្រីបពិសេសទេ។ ទោះយ៉ាងណាក៏ដោយ ពណ៌របស់វាត្រូវគ្នានឹងពណ៌ទូទៅនៃតំបន់ ឧទាហរណ៍ដូចជាព្រៃឈើ។ ជាឧទាហរណ៍ សត្វកណ្ដៀរជាច្រើនដែលមានស្លាបរបស់វាលាតសន្ធឹង អង្គុយលើសំបកឈើពណ៌ស ហើយមានស្លាបងងឹត ឈរយ៉ាងខ្លាំងប្រឆាំងនឹងផ្ទៃខាងក្រោយស្រាលនៃសំបកឈើ។ ទោះជាយ៉ាងណាក៏ដោយ ពួកវានៅតែមើលមិនឃើញ ព្រោះវាស្រដៀងនឹងចំណុចខ្មៅដែលអាចកើតមាននៅលើសំបកឈើនេះ។ ករណីបែបនេះគឺជារឿងធម្មតាណាស់។ អ្នកនិពន្ធនៃបន្ទាត់ទាំងនេះបានសង្កេតឃើញមេអំបៅ twilight មួយពីគ្រួសារ Notodontitae - Lophopteryx camelina ។ ដោយស្លាបរបស់វាបត់ មេអំបៅស្រដៀងនឹងសំបកឈើពណ៌លឿង។ មេអំបៅបានហោះចេញពីដើមឈើ ហើយ "ជាប់គាំង" នៅក្នុងម្ជុលស្រល់ ដែលមិនឆ្ងាយពីដី នៅតែគ្មានចលនា។ អាចមើលឃើញយ៉ាងច្បាស់នៅក្នុងបន្ទប់ទទួលភ្ញៀវពណ៌បៃតង វានៅតែមិនអាចមើលឃើញដោយសារតែវាស្រដៀងទៅនឹងបន្ទះពណ៌លឿង។ បានទម្លាក់ចូលទៅក្នុងសំណាញ់ វានៅតែស្ថិតក្នុងស្ថានភាពនៃ thanatosis ហើយភាពស្រដៀងគ្នារបស់វាទៅនឹងសំបកឈើបន្តធ្វើឱ្យមានការយល់ច្រឡំ។ បាតុភូតបែបនេះនៃភាពប្រហាក់ប្រហែលនឹងវត្ថុមួយក្នុងចំណោមវត្ថុដែលអាចធ្វើទៅបានក្នុងស្ថានភាពដែលបានផ្តល់ឱ្យអាចត្រូវបានគេហៅថាពណ៌មិនសំខាន់ពិសេស។
ពីករណីបែបនេះមនុស្សម្នាក់អាចរកឃើញការផ្លាស់ប្តូរជាច្រើនទៅភាពស្រដៀងគ្នាពិសេសបន្ថែមទៀត។
ជាឧទាហរណ៍ អាល់ប៊ុម Polygonium c-album របស់យើង ពេលអង្គុយនៅលើដីព្រៃ ក្លាយជាដូចជាស្លឹកស្ងួត។ មេអំបៅ Diphtera alpium អង្គុយលើសំបកឈើ ធ្វើត្រាប់តាមលំនាំ និងពណ៌នៃ lichen ។ល។
នៅក្នុងករណីទាំងនេះ សំណួរគឺអំពីពណ៌គ្រីបពិសេសជាង។

ដោយជ្រើសរើសស៊េរីនៃប្រភេទសត្វពីពណ៌ដែលមិនពិសេសទៅពណ៌គ្រីប យើងនឹងទទួលបានរូបភាពនៃការវិវត្តនៃបាតុភូតនេះ។ ទោះជាយ៉ាងណាក៏ដោយ ភាពជឿជាក់ជាងនេះទៅទៀតគឺការពិតដែលថាការកែលម្អលក្ខណៈសំខាន់ៗអាចត្រូវបានបញ្ជាក់នៅក្នុងប្រភេទសត្វមួយ។ ដូច្នេះ Schwanvin (1940) បានបង្ហាញថានៅក្នុងប្រភេទដូចគ្នានៃមេអំបៅ Zaretes isidora វាអាចធ្វើទៅបានដើម្បីបង្កើតទម្រង់ជាច្រើនដែលតួអក្សរគ្រីប (ស្រដៀងទៅនឹងស្លឹកស្ងួត) ឈានដល់កម្រិតនៃភាពល្អឥតខ្ចោះខុសៗគ្នា។ តួលេខនេះបង្ហាញពីទម្រង់ដើមនៃជម្ងឺ Zaretes isidora formaitis ។ ដូចដែលអ្នកអាចមើលឃើញ ឆ្នូតបណ្តោយ (ឡើងលើ) លាតសន្ធឹងតាមស្លាបខាងក្រោយ ធ្វើត្រាប់តាមផ្នែកកណ្តាលនៃស្លឹកស្ងួត។ ទោះយ៉ាងណាក៏ដោយ ការក្លែងបន្លំនេះនៅតែមិនល្អឥតខ្ចោះ។ ការបន្តនៃ "ពាក់កណ្តាល" នៃស្លឹកនៅក្នុងផ្នែកខាងមុខគឺនៅតែមិនច្បាស់លាស់ហើយលើសពីនេះទៀតឥទ្ធិពលគ្រីបត្រូវបានកាត់បន្ថយដោយវត្តមាននៃឆ្នូតផ្សេងទៀត (E3, Ua, E3p) ដែលរំខានដល់ភាពស្រដៀងគ្នាជាមួយផ្នែកកណ្តាលនៃស្លឹក។ . ទម្រង់មួយទៀតមាន Zaretes isidora f. strigosa - ភាពស្រដៀងនឹងស្លឹកគឺធំជាង។ សរសៃវ៉ែនកណ្តាល (ឡើង) គឺច្បាស់ជាងនេះទៅទៀត អ៊ី 3 បានបែកបាក់ដោយផ្នែក យូ ស្ថិតក្នុងស្ថានភាពនៃការបំផ្លិចបំផ្លាញទាំងស្រុង ដូច E 3 r ។ នៅលើផ្នែកខាងមុខ ឆ្អឹងជំនីរមានការរីកចម្រើនយ៉ាងខ្លាំង ហើយឆ្នូតងងឹតជាបន្តបន្ទាប់នៅមូលដ្ឋាននៃផ្នែកខាងមុខកំពុងរងការរិចរិល។ អរគុណចំពោះបញ្ហានេះ ប្រសិទ្ធភាពនៃការក្លែងធ្វើផ្នែកកណ្តាលនៃស្លឹកត្រូវបានពង្រឹង។ ប្រសិនបើឥឡូវនេះយើងប្រៀបធៀបមេអំបៅទាំងនេះជាមួយ callima យើងនឹងឃើញថាឥទ្ធិពលគ្រីបរបស់វាកាន់តែល្អឥតខ្ចោះ។ ដូច្នេះនៅក្នុង Zaretes ការបន្តនៃខ្សែដែលធ្វើត្រាប់តាមផ្នែកកណ្តាលនៃស្លឹកនៅលើស្លាបខាងមុខត្រូវបានផ្លាស់ប្តូរបន្តិច។ នេះមិនត្រូវបានគេសង្កេតឃើញនៅក្នុងកាលីម៉ាទេ។ ដូច្នេះ ដោយប្រើឧទាហរណ៍នៃទម្រង់ទាំងពីរ និង callima វាត្រូវបានបង្ហាញថាភាពស្រដៀងទៅនឹងស្លឹកមួយត្រូវបានសម្រេចយ៉ាងច្បាស់ដោយការផ្លាស់ទីលំនៅជាបន្តបន្ទាប់ និងបំផ្លាញផ្នែកទាំងអស់នៃការរចនាដែលបំពានឥទ្ធិពលគ្រីប។ ឧទាហរណ៍នេះបង្ហាញថាភាពស្រដៀងនឹងស្លឹកមិនបានកើតឡើងភ្លាមៗនោះទេ ប៉ុន្តែមានការអភិវឌ្ឍ និងប្រសើរឡើង។ លើសពីនេះទៅទៀតទម្រង់ទាំងពីរ - Zaretes forma itis និង f ។ strigosa គឺជាឧទាហរណ៍នៃកម្រិតខុសគ្នានៃឥទ្ធិពលដែលសម្រេចបាន។ បាតុភូតទាំងនេះគឺស្របទាំងស្រុងជាមួយនឹងទ្រឹស្ដីនៃការជ្រើសរើស ហើយជាភស្តុតាងដោយប្រយោលអំពីវា។
ទោះជាយ៉ាងណាក៏ដោយអ្វីដែលសំខាន់ជាងនេះទៅទៀតនោះគឺការពិតដែលថាផ្នែកកណ្តាលនៃស្លាប Callima បានកើតឡើងមួយផ្នែកដោយសារតែធាតុផ្សេងគ្នានៃគំរូជាងរបស់ Zaretes ។ ដូច្នេះមានផលដូចគ្នា។ ប្រភពដើមផ្សេងៗគ្នា. ការធ្វើត្រាប់តាមស្លឹកឈើត្រូវបានសម្រេចតាមវិធីផ្សេងៗគ្នា។ វាច្បាស់ណាស់ថាកត្តាដែលទទួលខុសត្រូវចំពោះលទ្ធផលទាំងនេះមិនមែនជាអាកាសធាតុ ឬលំហាត់ប្រាណនោះទេ ប៉ុន្តែជាភ្នែករបស់សត្វមំសាសី។ សត្វស្លាបបានសម្លាប់ចោលទម្រង់មិនសូវស្រដៀងនឹងស្លឹក ខណៈពេលដែលទម្រង់ស្រដៀងនឹងវានៅរស់រានមានជីវិត។
ចំពោះកត្តាផ្លូវចិត្តដែលសន្មត់ថាបណ្តាលឱ្យមានបាតុភូតដែលបានពិពណ៌នា ភស្តុតាងដ៏ល្អបំផុតដែលបដិសេធគំនិតមិនសមហេតុផលនេះគឺករណីនៃការធ្វើត្រាប់តាមរុក្ខជាតិ នៅពេលដែលឧទាហរណ៍ សត្វល្អិតធ្វើជាគំរូ ហើយផ្កាគឺជាអ្នកត្រាប់តាម។

រូបភាពបង្ហាញពីផ្កាអ័រគីដេ Ophrys muscifera ដែលមើលទៅគួរអោយកត់សំគាល់ដូចជាសត្វឃ្មុំ។ ភាពស្រដៀងគ្នានេះគឺផ្អែកលើ៖
1) ផ្កានេះត្រូវបានលំអងដោយសត្វល្អិត។ 2) ផ្កាមិនមានក្លិនទេ ហើយសត្វល្អិតដែលលំអងវាមិនស្វែងរកទឹកដម ហើយមិនទទួល។ 3) មានតែបុរសទេដែលមកទស្សនាផ្កា។ 4) ផ្កាក្នុងកម្រិតជាក់លាក់មួយ ស្រដៀងនឹងសត្វញីនៃប្រភេទសត្វល្អិតដូចគ្នា។ 5) បុរសអង្គុយលើផ្កាមានអាកប្បកិរិយាដូចគ្នានឹងពេលរួមរស់ជាមួយស្រី 6) ប្រសិនបើអ្នកដកផ្នែកខ្លះនៃផ្កាដែលធ្វើឱ្យវាស្រដៀងនឹងស្ត្រីនោះផ្កាមិនទាក់ទាញបុរសទេ (Kozo-Polyansky, ១៩៣៩)។ លក្ខណៈពិសេសទាំងអស់នេះបង្ហាញថា លក្ខណៈសម្ងាត់នៃផ្កាគឺជាការសម្របខ្លួនដ៏គួរឱ្យកត់សម្គាល់ចំពោះការលំអង។ ក្នុងករណីនេះ វាច្បាស់ណាស់ថា ទាំងទ្រឹស្តីនៃ "លំហាត់ប្រាណ" ឬឥទ្ធិពលនៃកត្តាអាកាសធាតុ និងផ្លូវចិត្តមិនពន្យល់អ្វីទាំងអស់។ ករណីដែលបានពិពណ៌នាគឺអាចយល់បានតែពីទស្សនៈនៃទ្រឹស្តីនៃការជ្រើសរើស ហើយជាភស្តុតាងប្រយោលដ៏ឆើតឆាយបំផុតមួយរបស់វា (Kozo-Polyansky, 1939) ។
ការសិក្សាអំពីច្បាប់ជាមូលដ្ឋាននៃការធ្វើត្រាប់តាមនាំឱ្យមានការសន្និដ្ឋានដូចគ្នា។ យើងធ្វើបទបង្ហាញអំពីគំរូសំខាន់ៗទាំងនេះ (Carpenter and Ford, 1936)។
ក) ការធ្វើត្រាប់តាម ប៉ះពាល់តែលក្ខណៈដែលអាចមើលឃើញ ឬហៅថាមុខងារដែលមើលឃើញប៉ុណ្ណោះ។
ខ) លក្ខណៈជាប្រព័ន្ធនៃគំរូ និងម៉ាស៊ីនក្លែងធ្វើអាចមាន ហើយជាក្បួនគឺខុសគ្នាទាំងស្រុង (ពោលគឺពួកវាជាកម្មសិទ្ធិរបស់ក្រុមប្រព័ន្ធខុសគ្នាទាំងស្រុង)។ ប៉ុន្តែនៅក្នុងរូបរាង (មើលឃើញ) ម៉ាស៊ីនក្លែងធ្វើគឺស្រដៀងនឹងម៉ូដែលមិនធម្មតា។
គ) ម៉ាស៊ីនក្លែងធ្វើ និងគំរូ ជាក្បួនកាន់កាប់តំបន់ចែកចាយដូចគ្នា។
ឃ) ម៉ាស៊ីនក្លែងធ្វើ និងគំរូហោះហើរជាមួយគ្នា។
ង) អ្នកក្លែងបន្លំខុសពីរូបរាងធម្មតា។ ក្រុមប្រព័ន្ធដែលគាត់ជាកម្មសិទ្ធិ។
គំរូទាំងនេះមិនអាចពន្យល់បានដោយលំហាត់ស្រដៀងគ្នានៃគំរូទេ។ ភាពមិនសមហេតុផលនៃ "ការពន្យល់" នេះគឺបង្ហាញឱ្យឃើញដោយខ្លួនឯង ជាពិសេសទាក់ទងនឹងការធ្វើត្រាប់តាមរុក្ខជាតិ។ ការពន្យល់នេះគឺមិនទំនងទាល់តែសោះក្នុងការទាក់ទងទៅនឹងសត្វល្អិត ដែលផ្តល់ឱ្យយ៉ាងជាក់លាក់ ចំនួនធំបំផុតឧទាហរណ៍នៃការធ្វើត្រាប់តាម។ ជាទូទៅ វាមិនអាចមានចម្ងល់អំពីសត្វនោះទេ ពោលគឺតិចជាងរុក្ខជាតិមួយ ដែលត្រាប់តាមរូបរាងរបស់វាដូចជាគំរូតាមរយៈការធ្វើលំហាត់ប្រាណ។ គេអាចសន្មត់ថា គំរូ និងម៉ាស៊ីនក្លែងធ្វើ ការរស់នៅជាមួយគ្នាត្រូវបានជះឥទ្ធិពលដោយកត្តាដូចគ្នា ដូច្នេះហើយគឺស្រដៀងគ្នា។
ទោះជាយ៉ាងណាក៏ដោយ វាប្រែថាអាហាររបស់គំរូ និងអ្នកត្រាប់តាម ក៏ដូចជាបរិយាកាសដែលពួកគេអភិវឌ្ឍ ជារឿយៗមានភាពខុសប្លែកគ្នាយ៉ាងខ្លាំង។ ដូច្នេះការពន្យល់សរីរវិទ្យានៃការធ្វើត្រាប់តាមមិនផ្តល់អ្វីទាំងអស់។ មានតែទ្រឹស្ដីនៃការជ្រើសរើសប៉ុណ្ណោះដែលអាចពន្យល់ពីការធ្វើត្រាប់តាម។ ដូចជាពណ៌សម្ងាត់ ការធ្វើត្រាប់តាមបានកើតឡើង និងត្រូវបានបង្កើតឡើងដោយសារតែឧបករណ៍ប្រើប្រាស់របស់វា។ ការទទួលបានលក្ខណៈត្រាប់តាម បង្កើនសក្តានុពលរស់រានមានជីវិត ហើយជាលទ្ធផល សក្តានុពលជីវសាស្ត្រនៃប្រភេទសត្វ។ ដូច្នេះហើយ ការជ្រើសរើសបានដើរក្នុងទិសដៅនៃការបង្កើតលក្ខណៈត្រាប់តាមតាមរយៈការបំផ្លាញអ្នកត្រាប់តាមដែលមិនសូវជោគជ័យ។ យើងនឹងឃើញនៅពេលក្រោយថាការសន្និដ្ឋាននេះត្រូវបានបញ្ជាក់ដោយពិសោធន៍។
4. ពណ៌និងរូបរាង Aposematic. ពីការបង្ហាញមុនវាច្បាស់ណាស់ថាមូលដ្ឋាននៃបាតុភូតនៃការធ្វើត្រាប់តាមគឺភាពស្រដៀងគ្នានៃអ្នកត្រាប់តាមគំរូ។ ភាពស្រដៀងគ្នានេះគឺផ្អែកលើការពិតដែលថាឧទាហរណ៍គំរូគឺមិនអាចបរិភោគបានហើយដូច្នេះភាពស្រដៀងគ្នាទៅនឹងវាបញ្ឆោតសត្រូវដែល "ធ្វើខុស" សត្វល្អិតដែលអាចបរិភោគបានសម្រាប់សត្វដែលមិនអាចបរិភោគបាន។ ដូច្នេះនៅក្នុងប្រភពដើមរបស់ពួកគេ ប្រភេទសត្វត្រាប់តាមគឺទាក់ទងយ៉ាងច្បាស់ទៅនឹងប្រភេទសត្វគំរូ។ ភាពមិនអាចបរិភោគបានគឺដោយសារតែក្លិនមិនល្អ សារធាតុពុល ឬការដុតនៃសារធាតុសំងាត់ សរីរាង្គដែលមានក្លិនស្អុយ។ល។ លក្ខណៈសម្បត្តិទាំងនេះជាធម្មតាត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងពណ៌ភ្លឺ និងគួរឱ្យកត់សម្គាល់ លំនាំមុតស្រួច ឧទាហរណ៍ ឆ្នូតពណ៌លឿងងងឹត និងភ្លឺឆ្លាស់គ្នា ដូចដែលយើងឃើញនៅក្នុងសត្វស្វា។ ឬផ្ទៃខាងក្រោយពណ៌ក្រហម ឬលឿងភ្លឺដែលមានចំណុចខ្មៅ (ដូចជា សត្វខ្លាឃ្មុំ) ជាដើម សត្វដង្កូវស៊ីរបស់មេអំបៅជាច្រើនមានពណ៌ភ្លឺខ្លាំង និងមានពណ៌ចម្រុះ។ ជាមួយនឹងពណ៌ និងការរចនាដ៏ភ្លឺទាំងនេះ សត្វល្អិតហាក់ដូចជា "ប្រកាស" ភាពមិនអាចទទួលយកបានរបស់វា ឧទាហរណ៍ សត្វស្លាបរៀនពីបទពិសោធន៍ផ្ទាល់ខ្លួនដើម្បីសម្គាល់សត្វល្អិតបែបនេះ ហើយជាក្បួនកុំប៉ះពួកវា។ ពីនេះវាកាន់តែច្បាស់ថាភាពស្រដៀងទៅនឹងសត្វល្អិតដែលមិនអាចបរិភោគបានបែបនេះមានអត្ថន័យដ៏មានប្រយោជន៍ និងដើរតួរនាទីនៃការសម្របខ្លួនដែលមើលឃើញដែលវិវឌ្ឍន៍នៅក្នុងសត្វល្អិតដែលអាចបរិភោគបាន។ នេះគឺជាកន្លែងដែលបាតុភូតនៃការធ្វើត្រាប់តាមកើតឡើង។ យើងនឹងឃើញនៅពេលក្រោយដែលការពន្យល់អំពីការធ្វើត្រាប់តាមនេះត្រូវបានបញ្ជាក់ដោយពិសោធន៍។ ពណ៌ និងលំនាំព្រមានត្រូវបានគេហៅថា aposematic ហើយលំនាំធ្វើត្រាប់តាមដែលត្រូវគ្នាត្រូវបានគេហៅថា pseudoaposematic ។
5. ទីបំផុតអនុញ្ញាតឱ្យយើងរស់នៅលើបាតុភូតនេះ។ ពណ៌ទទួលស្គាល់ជួនកាលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងអាកប្បកិរិយាដែលត្រូវគ្នា។ ឧទាហរណ៍មួយគឺការទទួលស្គាល់ពណ៌នៃ moorhen (Zhitkov និង Buturlin, 1916) ។ ពណ៌រោមរបស់បក្សីនេះគឺសម្ងាត់។ មានតែផ្នែកខាងក្រោមប៉ុណ្ណោះដែលត្រូវបានលាបពណ៌ឱ្យស្អាត ពណ៌ស. Moorhen ជាប់នឹងវាលភក់ដ៏ក្រាស់។ កូនរបស់បក្សីនេះមានកូនមាន់ប្រហែល 12 ក្បាល។ វាពិបាកណាស់ក្នុងការរក្សាកូនមាន់មួយក្រុមនេះឱ្យនៅជាមួយគ្នាក្នុងព្រៃក្រាស់។ បក្សីអាចគេចចេញពីម្តាយបានយ៉ាងងាយ បាត់បង់ការមើលឃើញរបស់នាង និងក្លាយជាសត្វព្រៃ សត្វមំសាសីតូច. ដូច្នេះហើយ សត្វមូរដែលធ្វើដំណើរឆ្លងកាត់ព្រៃក្រាស់ លើកកន្ទុយរបស់នាងឱ្យខ្ពស់ លាតត្រដាងផ្នែកខាងក្រោមពណ៌ស ដែលដើរតួជា "សញ្ញាណែនាំ" សម្រាប់កូនមាន់ ដែលដឹកនាំដោយពួកគេដើរតាមម្តាយរបស់ពួកគេដោយមិននឹកស្មានដល់។
ដូច្នេះ កន្ទុយពណ៌សនៃ moorhen គឺជាការសម្របខ្លួនដែលបង្កើនអត្រារស់រានមានជីវិតរបស់កូនចៅ។
ករណីដែលបានពិពណ៌នាគឺគួរឱ្យចាប់អារម្មណ៍ទោះជាយ៉ាងណាពីផ្នែកផ្សេងទៀត។ សត្វស្លាបជាច្រើនមានពណ៌ស ហើយវាប្រហែលជាមិនមានអត្ថន័យដូចខាងលើទេ។ ការកត់សម្គាល់ស្រដៀងគ្នានេះត្រូវបានធ្វើឡើងដោយពួកប្រឆាំងដាវីន ដែលបានចង្អុលបង្ហាញថាលក្ខណៈមួយកើតឡើងដោយមិនគិតពីប្រយោជន៍របស់វា។
ទោះជាយ៉ាងណាក៏ដោយ ការកត់សម្គាល់នេះគ្រាន់តែជាភស្តុតាងនៃការយល់ច្រឡំនៃទ្រឹស្តីនៃការជ្រើសរើសប៉ុណ្ណោះ។ លក្ខណៈមួយក្លាយជាការសម្របខ្លួនតែនៅក្រោមលក្ខខណ្ឌនៃទំនាក់ទំនងជាក់លាក់ជាមួយស្ថានភាពជីវិតជុំវិញ។ នៅក្នុងលក្ខខណ្ឌផ្សេងទៀតគាត់អាចព្រងើយកណ្តើយ។ ដូច្នេះឧទាហរណ៍ដែលបានវិភាគគឺជាភស្តុតាងបន្ថែមទៀតនៃការពិតដែលថាការសម្របខ្លួនមិនមែនជាអ្វីដែលដាច់ខាតនោះទេប៉ុន្តែគ្រាន់តែជាបាតុភូតនៃទំនាក់ទំនងនៃលក្ខណៈដែលបានផ្តល់ឱ្យទៅនឹងលក្ខខណ្ឌបរិស្ថានជាក់លាក់។
6. លក្ខណៈពិសេសនៃតួអក្សរប្រែប្រួលនៃផ្កា entomo-, ornitho- និង theriophilous. យើងបានពណ៌នារួចហើយអំពីការសម្របខ្លួននៃផ្កា entomophilous ទៅនឹង pollination ដោយសត្វល្អិត។ ការលេចឡើងនៃការសម្របខ្លួនទាំងនេះនៅក្រោមឥទ្ធិពលនៃការជ្រើសរើសគឺបង្ហាញឱ្យឃើញដោយខ្លួនឯងព្រោះវាមិនអាចទៅរួចទេក្នុងការពន្យល់ពីការសម្របខ្លួននៃផ្កា entomophilous ទៅនឹងសត្វល្អិតដោយទ្រឹស្តីផ្សេងទៀតណាមួយ។
ឧទាហរណ៍មិនគួរឱ្យចាប់អារម្មណ៍តិចជាងនៃសកម្មភាពនៃការជ្រើសរើសគឺជាតួអក្សរសម្របខ្លួននៃផ្កា ornitho និង theriophilous ។
ផ្កា Ornithophilous ត្រូវបានសម្របខ្លួនទៅនឹងលំអងដោយបក្សី។ បក្សីរុករកដោយការមើលឃើញ។ ផ្កាគួរតែមានពណ៌ភ្លឺខណៈពេលដែលក្លិនមិនសំខាន់។ ដូច្នេះផ្កា ornithophilous ជាក្បួនមិនមានក្លិនទេ។ ទោះជាយ៉ាងណាក៏ដោយពួកគេមានពណ៌ភ្លឺដែលទាក់ទាញសត្វស្លាប។ ឧទាហរណ៍ ផ្កាលំអងដោយសត្វស្លាបមានពណ៌ក្រហម ខៀវ ឬបៃតង ដែលត្រូវនឹងពណ៌សុទ្ធនៃវិសាលគមព្រះអាទិត្យ។ ប្រសិនបើនៅក្នុងក្រុមដូចគ្នានៃរុក្ខជាតិមានទម្រង់ ornithophilous នោះពួកវាមានពណ៌នៃវិសាលគមខណៈពេលដែលរុក្ខជាតិផ្សេងទៀតមិនមានពណ៌ស្រដៀងគ្នា។ ដូច្នេះវាច្បាស់ណាស់ថាពណ៌ផ្កា ornithophilous គឺជាការសម្របខ្លួនទៅនឹងការមកទស្សនាដោយសត្វស្លាប។ ទោះជាយ៉ាងណាក៏ដោយអ្វីដែលគួរអោយកត់សំគាល់បំផុតនោះគឺថាផ្កា ornithophilous ត្រូវបានសម្របខ្លួនទៅនឹងសត្វស្លាបមិនត្រឹមតែពណ៌ប៉ុណ្ណោះទេប៉ុន្តែថែមទាំងនៅក្នុងរចនាសម្ព័ន្ធរបស់វាផងដែរ។ ដូច្នេះពួកវាជួបប្រទះការកើនឡើងនៃកម្លាំងនៃផ្កាដោយសារតែការអភិវឌ្ឍនៃជាលិកាមេកានិច (នៅក្នុង xerophytes) ឬការកើនឡើងនៃ turgor (នៅក្នុងរុក្ខជាតិនៃតំបន់ត្រូពិចសើម) ។ ផ្កា Ornithophilous បញ្ចេញសារធាតុរាវច្រើនក្រៃលែង ឬទឹកដមស្អិត។
ផ្ការបស់រុក្ខជាតិ ornithophilous Holmskioldia sanguinea មានកាលីបរាងពងក្រពើ។ វាកើតឡើងពីការលាយបញ្ចូលគ្នានៃសរីរាង្គស្លឹកទាំងប្រាំ ហើយមានរាងដូចចីវលោមដែលមានពណ៌ក្រហមដ៏កាចសាហាវ។ ផ្កាកូរ៉ូឡាមានពណ៌ដូចគ្នា មានរាងដូចស្នែងម៉ាញ់។ ដើមមានរាងកោង ហើយចេញទៅខាងក្រៅ ដូចជាដុំពក។ ផ្កាគឺគ្មានក្លិន; ការបញ្ចេញទឹកដមដ៏អស្ចារ្យបំផុតកើតឡើងនៅពេលព្រឹកព្រលឹមក្នុងអំឡុងម៉ោងហោះហើរនៃសត្វស្លាបព្រះអាទិត្យ Cirnirys pectoralis ។ សត្វស្លាបទម្លាក់ចំពុះកោងរបស់ពួកគេចូលទៅក្នុងផ្កាកូរ៉ូឡា អង្គុយលើផ្កា ឬឈប់នៅពីមុខវានៅលើអាកាស ដូចជាសត្វស្លាបហើរ ពោលគឺហើរស្លាប។ ចំពុះត្រូវគ្នានឹងខ្សែកោងនៃផ្កាកូរ៉ូឡា។ ចំពុះនេះហាក់ដូចជាមានរាងដូចផ្កាកូរ៉ូឡា ហើយចំពុះនោះដូចជារបាំងមុខរបស់បក្សី។ នៅពេលដែលចំពុះត្រូវលិចទឹក អន្ទង់ប៉ះនឹងរោមថ្ងាស ហើយលំអងវា។ នៅពេលទៅទស្សនាផ្កាមួយផ្សេងទៀត លំអងបានយ៉ាងងាយធ្លាក់លើការមាក់ងាយ ហើយការលំអងឆ្លងកាត់កើតឡើង (Porsch, 1924) ។
ជាចុងក្រោយ ចូរយើងរស់នៅលើផ្កាដែលអាចត្រូវបានគេហៅថា theriophilic ពោលគឺប្រែប្រួលទៅនឹងការលំអងដោយថនិកសត្វ ជាពិសេសសត្វប្រចៀវ។ ផ្កា Theriophilous មានលក្ខណៈពិសេសមួយចំនួន។ សត្វប្រចៀវអាចបំផ្លាញផ្កាយ៉ាងងាយស្រួល។ ក្នុងន័យនេះ ផ្កា theriophilous ដែលសម្របទៅនឹងការលំអងដោយសត្វប្រចៀវត្រូវបានសម្គាល់ដោយភាពខ្លាំងមិនធម្មតានៃជាលិការបស់ពួកគេ ហើយផ្នែកនីមួយៗរបស់ពួកគេ (ដូចនៅក្នុងករណីនៃផ្កាអ័រនីហ្វីល) ត្រូវបានផ្សំជាមួយគ្នា។ ចាប់តាំងពីសត្វប្រចៀវហើរនៅពេលព្រលប់ ផ្កា theriophilous បញ្ចេញក្លិនក្រអូបតែនៅពេលនេះ។ ក្នុងពេលព្រលប់ពួកគេក៏លួចបញ្ចេញទឹកដម (Porsch)។ សម្រាប់ផ្នែករបស់ពួកគេ សត្វប្រចៀវខ្លះដែលប្រើផ្កាត្រូវបានសម្របតាមក្រោយ។ ដូច្នេះ បិសាចអណ្តាតវែង (Glossophaga soricina) មានមាត់វែង ហើយអណ្តាតត្រូវបានពន្លូត និងបំពាក់ដោយជក់ដែលប្រមូលទឹកដម។
ដូច្នេះរចនាសម្ព័ននិងពណ៌នៃផ្កាធម្មជាតិនៃក្លិនឬអវត្តមានរបស់វាក៏ដូចជាពេលវេលានៃការបញ្ចេញទឹកដមប្រែជាប្រែប្រួលជាមួយនឹងភាពត្រឹមត្រូវដ៏អស្ចារ្យចំពោះអ្នកទស្សនា (មេអំបៅសត្វរុយសត្វបក្សីថនិកសត្វ) ដែលត្រូវគ្នានឹងពួកគេ។ អង្គការ ពេលវេលាហោះហើរ និងចរិតលក្ខណៈ។
ស្ទើរតែគ្មានតម្រូវការដើម្បីបញ្ជាក់ថាបើគ្មានទ្រឹស្ដីនៃការជ្រើសរើសទេ ការសម្របខ្លួនដែលបានពិពណ៌នាទាំងអស់នឹងត្រូវសន្មតថាជា "សមត្ថភាព" ដែលមិនអាចយល់បាន និងអាថ៌កំបាំងទាំងស្រុងក្នុងការទទួលបានរចនាសម្ព័ន្ធសមស្រប សម្របតាមគ្រប់លម្អិតសម្រាប់អ្នកទស្សនាផ្កា។ ផ្ទុយទៅវិញ ទ្រឹស្តីនៃការជ្រើសរើសផ្តល់នូវការពន្យល់ធម្មជាតិទាំងស្រុងសម្រាប់បាតុភូតដែលបានពិពណ៌នា។ ទង្វើនៃការលំអងឈើឆ្កាងគឺជាគុណភាពដ៏សំខាន់មួយ ដោយគ្មានការបន្តពូជរបស់កូនចៅគឺពិបាកណាស់។ ដូច្នេះ រុក្ខជាតិដែលសម្របតាមភ្នាក់ងារលម្អងរបស់វាកាន់តែល្អ ឱកាសបន្តពូជក៏កាន់តែធំ។
ដូច្នេះ ការសម្របខ្លួនត្រូវបានគោរពដោយជៀសមិនរួចចំពោះកម្រិតនៃភាពល្អឥតខ្ចោះខ្ពស់ ដែលពួកវាចាំបាច់សម្រាប់ជីវសាស្រ្ត។
វាគួរឱ្យកត់សម្គាល់ថាភាពល្អឥតខ្ចោះនិងភាពជាក់លាក់នៃការសម្របខ្លួននេះគឺខ្ពស់ជាពិសេសនៅពេលដែលផ្កាត្រូវបានទៅទស្សនាដោយអ្នកប្រើប្រាស់ជាក់លាក់នៃទឹកដម។ ប្រសិនបើនេះមិនមែនជាករណីទេនោះការសម្របខ្លួនទៅនឹងពួកគេជាក្បួនមានលក្ខណៈទូទៅជាង។
7. ឥឡូវនេះ ចូរយើងរស់នៅលើកោះដែលមិនមានស្លាបជាឧទាហរណ៍នៃភស្តុតាងដោយប្រយោលនៃការជ្រើសរើសធម្មជាតិ។ ដោយសំដៅទៅលើ Wollaston លោក Darwin បានចង្អុលទៅការពិតដែលថានៅលើ Fr. ក្នុងចំណោម 550 ប្រភេទនៃ Madera beetles, 200 ប្រភេទគឺមិនអាចហោះហើរបាន។ បាតុភូតនេះត្រូវបានអមដោយរោគសញ្ញាដូចខាងក្រោម។ ការពិតមួយចំនួនបង្ហាញថា សត្វកកេរហើរជាញឹកញាប់ត្រូវបានខ្យល់បក់ចូលទៅក្នុងសមុទ្រ ហើយស្លាប់។ ម្យ៉ាងវិញទៀត Wollaston បានកត់សម្គាល់ឃើញថា សត្វឃ្មុំ Madeira លាក់ខ្លួនខណៈពេលដែលខ្យល់បក់មក ហើយគ្មានព្រះអាទិត្យ។ លើសពីនេះ វាត្រូវបានបញ្ជាក់ថា សត្វល្អិតគ្មានស្លាប គឺជាលក្ខណៈពិសេសនៃកោះដែលមានខ្យល់បក់ខ្លាំង។ ពីការពិតទាំងនេះ Darwin បានសន្និដ្ឋានថាភាពគ្មានការហោះហើរនៃសត្វល្អិតនៅលើកោះបែបនេះត្រូវបានបង្កើតឡើងដោយការជ្រើសរើស។ ទម្រង់ហោះហើរត្រូវបានគេយកទៅឆ្ងាយដោយខ្យល់ ហើយស្លាប់ ចំណែកទម្រង់គ្មានស្លាបត្រូវបានរក្សាទុក។ អាស្រ័យហេតុនេះ តាមរយៈការលុបបំបាត់ឥតឈប់ឈរនៃទម្រង់ស្លាប សត្វស្លាបនៃកោះមហាសមុទ្រដែលបក់បោកដោយខ្យល់ត្រូវបានបង្កើតឡើង។
ការសន្មត់ទាំងនេះត្រូវបានបញ្ជាក់ទាំងស្រុង។ វាត្រូវបានគេរកឃើញថាភាគរយនៃទម្រង់គ្មានស្លាបនៅលើកោះដែលមានខ្យល់បក់គឺតែងតែខ្ពស់ជាងនៅលើទ្វីប។ ដូច្នេះនៅលើកោះ Croceti ក្នុងចំណោម 17 ប្រភេទនៃសត្វល្អិត 14 គឺគ្មានស្លាប។ នៅលើកោះ Kerguelen ក្នុងចំណោមសត្វរុយសរុបចំនួនប្រាំបីប្រភេទ មានតែមួយប្រភេទប៉ុណ្ណោះដែលមានស្លាប។
ប្រាកដណាស់ គេអាចនិយាយបានថា ការជ្រើសរើសមិនមានជាប់ពាក់ព័ន្ធជាមួយវាទេ។ ជាឧទាហរណ៍ ពពួកសត្វដែលមិនមានស្លាបត្រូវបានគេសង្កេតឃើញនៅក្នុង Drosophila ។ អាស្រ័យហេតុនេះ ភាពគ្មានការហោះហើរគឺជាលទ្ធផលនៃការផ្លាស់ប្តូរ ហើយការជ្រើសរើសគ្រាន់តែ "យក" ការផ្លាស់ប្តូរ ប្រសិនបើវាមានប្រយោជន៍ ដូចករណីនៅលើកោះដែលមានខ្យល់បក់បោក។ ទោះជាយ៉ាងណាក៏ដោយ វាច្បាស់ណាស់ថាគ្មានស្លាបរបស់សត្វល្អិតកោះ ដែលបង្ហាញយ៉ាងច្បាស់ពីតួនាទីច្នៃប្រឌិតនៃការជ្រើសរើស។ ចូរយើងពិចារណាឧទាហរណ៍ដែលត្រូវគ្នា។
មួយក្នុងចំនោមសត្វរុយគ្មានស្លាប Kerguelen មានលក្ខណៈពិសេសមួយទៀតក្រៅពីគ្មានស្លាប៖ វាតែងតែនៅខាងក្រោមស្លឹករបស់រុក្ខជាតិដែលធន់នឹងខ្យល់។ លើសពីនេះទៅទៀត ជើងរបស់សត្វរុយនេះត្រូវបានបំពាក់ដោយក្រញ៉ាំជើងដ៏រឹងមាំ។ នៅក្នុងការហោះហើរ Kerguelen មួយផ្សេងទៀត - Amalopteryx maritima - រួមជាមួយនឹងការរីកដុះដាលនៃស្លាប ភ្លៅនៃជើង hind មានសាច់ដុំអភិវឌ្ឍខ្លាំងដែលត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងសមត្ថភាពលោតរបស់សត្វរុយ។ លើសពីនេះទៀតសត្វល្អិតទាំងនេះត្រូវបានកំណត់ដោយអាកប្បកិរិយាគួរឱ្យចាប់អារម្មណ៍។ ដរាបណាព្រះអាទិត្យគ្របដណ្ដប់ដោយពពក (ខ្យល់បក់) សត្វល្អិតដែលមិនចេះហោះហើរក៏លាក់ខ្លួនភ្លាមៗ ចូលទៅក្នុងដី លាក់ខ្លួនក្នុងព្រៃស្មៅក្រាស់ រំកិលទៅក្រោមស្លឹក។ល។ ត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងលក្ខណៈពិសេសមួយចំនួនផ្សេងទៀតនៃអង្គការ និងអាកប្បកិរិយា។ វាងាយស្រួលក្នុងការមើលឃើញភាពមិនអាចកាត់ថ្លៃបាននៃគុណភាព "កោះ" បែបនេះនៅក្នុងការផ្លាស់ប្តូរមួយ។ សំណួរគឺអំពីការប្រមូលផ្តុំ តាមរយៈសកម្មភាពនៃការជ្រើសរើស នៃភាពស្មុគស្មាញទាំងមូលនៃលក្ខណៈ "កោះ" ។
ភស្តុតាងប្រយោលដ៏គួរឲ្យកត់សម្គាល់បំផុតមួយនៃការជ្រើសរើសធម្មជាតិគឺលក្ខណៈនៃរុក្ខជាតិផ្កា Kerguelen ។ មិនមានរុក្ខជាតិលម្អងសត្វល្អិតនៅលើកោះទាំងនេះទេ។ ការពិតនេះនឹងកាន់តែច្បាស់ប្រសិនបើយើងចងចាំថាការហោះហើរត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការស្លាប់។ ដូច្នេះហើយ នៅលើកោះ Kerguelen ដែលបក់ដោយខ្យល់ មានតែរុក្ខជាតិលំអងខ្យល់ប៉ុណ្ណោះ។ វាច្បាស់ណាស់ថារុក្ខជាតិលម្អងដោយសត្វល្អិតមិនអាចរស់បាននៅលើកោះដោយសារតែកង្វះសត្វល្អិតដែលត្រូវគ្នា។ ក្នុងន័យនេះ រុក្ខជាតិផ្កា Kerguelen ក៏បាត់បង់ការសម្របខ្លួនទៅនឹងការបំពុលដោយសត្វល្អិត ជាពិសេសពណ៌ភ្លឺរបស់វា។ ឧទាហរណ៍នៅក្នុងផ្កាក្លាំពូ (Lyallia, Colobanthus) ផ្កាគឺគ្មានពណ៌ភ្លឺទេ ហើយនៅក្នុងផ្កាខាត់ណាខៀវក្នុងស្រុក (Ranunculus crassipes, R. trullifolius) ផ្កាត្រូវបានកាត់បន្ថយទៅជាឆ្នូតតូចចង្អៀត។ សម្រាប់ហេតុផលដែលបានរៀបរាប់ខាងលើ រុក្ខជាតិនៃកោះ Kerguelen មានភាពទាក់ទាញនៅក្នុងភាពក្រីក្រនៃពណ៌របស់វា ហើយយោងទៅតាមអ្នកធម្មជាតិម្នាក់ដែលបានសង្កេតឃើញវាបានទទួល "ពណ៌ស្រទន់" ។ បាតុភូតទាំងនេះបង្ហាញពីសកម្មភាពនៃការជ្រើសរើសធម្មជាតិជាមួយនឹងភាពច្បាស់លាស់មិនធម្មតា។
8. អាកប្បកិរិយាសម្របខ្លួនជាភស្តុតាងប្រយោលនៃការជ្រើសរើស. អាកប្បកិរិយារបស់សត្វនៅក្នុងករណីជាច្រើនបង្ហាញយ៉ាងច្បាស់ថាវាបានអភិវឌ្ឍនៅក្រោមឥទ្ធិពលនៃការជ្រើសរើស។ Kaftanovsky (1938) ចង្អុលបង្ហាញថា guillemots ពងរបស់ពួកគេនៅលើ cornices ដែលមានប្រជាជនយ៉ាងក្រាស់ដោយ guillemots ផ្សេងទៀត។ ការប្រយុទ្ធគ្នាយ៉ាងសាហាវកើតឡើងរវាងសត្វស្លាបនៅលើកន្លែងនីមួយៗ។ សត្វស្លាបផ្សេងទៀតស្វាគមន៍ ហ្គីលម៉ុត ដែលទើបនឹងមកដល់ថ្មី ជាមួយនឹងការវាយប្រហារដ៏រសើបពីចំពុះដ៏រឹងមាំរបស់វា។ យ៉ាងណាក៏ដោយ guillemot រឹងរូសប្រកាន់ខ្ជាប់នឹង cornices ដែលមានប្រជាជនច្រើនទាំងនេះ ទោះបីជាមានកន្លែងទំនេរនៅក្បែរនោះក៏ដោយ។ ហេតុផលសម្រាប់អាកប្បកិរិយានេះត្រូវបានពន្យល់យ៉ាងសាមញ្ញ។ Kaftanovsky ចង្អុលបង្ហាញថា ការសាយភាយ ពោលគឺ អាណានិគមដែលមានប្រជាជនតិចគឺទទួលរងការវាយប្រហារដោយសត្វឈ្មោល ខណៈពេលដែលអាណានិគមដែលមានប្រជាជនច្រើនមិនត្រូវបានវាយប្រហារដោយក្រុមក្រោយៗទៀត ឬងាយត្រូវបានបណ្តេញចេញដោយការវាយប្រហាររួម។
វាច្បាស់ណាស់អំពីរបៀបដែលសភាវគតិនៃអាណានិគមត្រូវបានបង្កើតឡើងក្នុងចំណោម guillemots ។ បុគ្គលដែលមិនមានសភាវគតិបែបនេះ ត្រូវទទួលរងការលុបបំបាត់ជាបន្តបន្ទាប់ ហើយស្ថានភាពអំណោយផលបំផុតគឺសម្រាប់បុគ្គលដែលស្វែងរកពងនៅក្នុងបរិយាកាសនៃអាណានិគមបក្សីដែលមានប្រជាជនច្រើន។
ការបង្ហាញជាពិសេសគឺជាឧទាហរណ៍នៃអាកប្បកិរិយាសម្របខ្លួនដែលទាក់ទងនឹងសកម្មភាពសភាវគតិសុទ្ធសាធឧទាហរណ៍នៅក្នុងសត្វល្អិត។ នេះរួមបញ្ចូលទាំងសកម្មភាពរបស់ Hymenoptera ជាច្រើន រួមទាំងសត្វខ្វិនមួយចំនួនដែលត្រូវបានពិពណ៌នាដោយ Fabre និងអ្នកស្រាវជ្រាវផ្សេងទៀត។ សត្វស្វាខ្លះវាយប្រហារ ឧទាហរណ៍ ពីងពាង ប្រើចង្កឹះរបស់វា ដើម្បីឆ្លងទៅមជ្ឈមណ្ឌលសរសៃប្រសាទ ហើយដាក់ពងនៅលើដងខ្លួនរបស់សត្វពីងពាង។ ដង្កូវញាស់ ស៊ីចំណីរស់ ប៉ុន្តែខ្វិន។ សត្វពីងពាងដែលខ្វិនសត្វពីងពាងវាយប្រហារមជ្ឈមណ្ឌលសរសៃប្រសាទរបស់វាដោយភាពច្របូកច្របល់ ហើយម្យ៉ាងវិញទៀត សត្វពីងពាងដែលឈ្លានពានចំពោះសត្វល្អិតផ្សេងទៀត ប្រែទៅជាអស់សង្ឃឹមប្រឆាំងនឹងប្រភេទនៃ wasp ដែលជាសត្រូវជាក់លាក់របស់វា។ មួយគូនៃប្រភេទជាក់លាក់មួយ - wasp និងពីងពាង ដែលជាមំសាសីខ្វិន និងសត្វព្រៃរបស់វា ដូច្នេះហើយ វាមានលក្ខណៈប្រែប្រួលទៅនឹងគ្នាទៅវិញទៅមក។ សត្វពីងពាងវាយប្រហារតែប្រភេទសត្វពីងពាងប៉ុណ្ណោះ ហើយសត្វពីងពាងគ្មានការការពារប្រឆាំងនឹងប្រភេទសត្វស្វាមួយប្រភេទ។ វាច្បាស់ណាស់ថាការបង្កើតការតភ្ជាប់ថេរបែបនេះរវាងប្រភេទជាក់លាក់ពីរអាចត្រូវបានពន្យល់តែលើមូលដ្ឋាននៃទ្រឹស្តីនៃការជ្រើសរើសប៉ុណ្ណោះ។ សំណួរគឺអំពីទំនាក់ទំនងដែលលេចចេញជាប្រវត្តិសាស្ត្ររវាងទម្រង់ដែលសមស្របបំផុតចំពោះគ្នាទៅវិញទៅមកនៅក្នុងទំនាក់ទំនងដែលបានពិពណ៌នា។
ចូរបន្តទៅភស្តុតាងផ្ទាល់នៃអត្ថិភាពនៃការជ្រើសរើសធម្មជាតិនៅក្នុងធម្មជាតិ។
ភស្តុតាងផ្ទាល់នៃការជ្រើសរើសធម្មជាតិ
ចំនួនភស្តុតាងដោយផ្ទាល់នៃការជ្រើសរើសធម្មជាតិត្រូវបានទទួលតាមរយៈការសង្កេតតាមវាលសមស្រប។ ក្នុងចំណោមការពិតមួយចំនួនធំ យើងនឹងលើកយកតែមួយចំនួនប៉ុណ្ណោះ។
1. ក្នុងអំឡុងពេលព្យុះនៅ New England សត្វចាបចំនួន 136 ក្បាលបានស្លាប់។ Bumpes (1899) បានពិនិត្យមើលប្រវែងនៃស្លាប កន្ទុយ និងចំពុះ ហើយវាបានប្រែក្លាយថាការស្លាប់គឺជាការជ្រើសរើស។ ភាគរយដ៏ធំបំផុតនៃអ្នកដែលត្រូវបានសម្លាប់គឺចាប ដែលត្រូវបានសម្គាល់ដោយស្លាបវែងជាងទម្រង់ធម្មតា ឬផ្ទុយទៅវិញ ស្លាបខ្លីជាង។ ដូច្នេះវាប្រែថាក្នុងករណីនេះមានការជ្រើសរើសសម្រាប់បទដ្ឋានជាមធ្យមខណៈពេលដែលទម្រង់គេចវេះបានស្លាប់។ នៅទីនេះយើងឃើញសកម្មភាពនៃការជ្រើសរើសដោយផ្អែកលើវិសមភាពរបស់បុគ្គលទាក់ទងនឹងកត្តាលុបបំបាត់ - ព្យុះ។
2. Weldon (1898) បានបង្កើតការពិតនៃលំដាប់ផ្ទុយ - ការរស់រានមានជីវិតនៃទម្រង់ intraspecific មួយនៅក្រោមលក្ខខណ្ឌធម្មតា និងការរស់រានមានជីវិតរបស់មួយផ្សេងទៀតនៅក្រោមលក្ខខណ្ឌផ្លាស់ប្តូរ។ Weldon បានសិក្សាពីភាពប្រែប្រួលនៃក្តាមមួយដែលក្នុងនោះមានទំនាក់ទំនងជាក់លាក់រវាងទទឹងនៃថ្ងាស និងប្រវែងនៃរាងកាយ ដែលត្រូវបានបង្ហាញនៅក្នុងការពិតដែលថានៅពេលដែលប្រវែងនៃរាងកាយផ្លាស់ប្តូរទទឹងនៃថ្ងាសក៏ផ្លាស់ប្តូរផងដែរ។ . វាត្រូវបានគេរកឃើញថានៅចន្លោះឆ្នាំ 1803 និង 1898 ទទឹងថ្ងាសជាមធ្យមនៃក្តាមមានប្រវែងជាក់លាក់មួយបានថយចុះជាលំដាប់។ Weldon បានបង្កើតឡើងថាការផ្លាស់ប្តូរនេះត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការផ្លាស់ប្តូរការសម្របខ្លួនអាស្រ័យលើការកើតឡើងនៃលក្ខខណ្ឌថ្មីនៃអត្ថិភាព។ នៅ Plymouth ជាកន្លែងដែលការសង្កេតត្រូវបានធ្វើឡើង ផែមួយត្រូវបានសាងសង់ដែលធ្វើឱ្យឥទ្ធិពលនៃជំនោរចុះខ្សោយ។ ជាលទ្ធផល បាតសមុទ្រនៃឆ្នេរសមុទ្រ Plymouth បានចាប់ផ្តើមកកស្ទះយ៉ាងខ្លាំងជាមួយនឹងភាគល្អិតដីដែលនាំមកដោយទន្លេ និងកាកសំណល់ទឹកស្អុយសរីរាង្គ។ ការផ្លាស់ប្តូរទាំងនេះបានប៉ះពាល់ដល់សត្វបាត ហើយ Weldon បានភ្ជាប់ពួកវាជាមួយនឹងការផ្លាស់ប្តូរទទឹងថ្ងាសរបស់ក្តាម។ ដើម្បីសាកល្បងនេះ ការពិសោធន៍ខាងក្រោមត្រូវបានអនុវត្ត។ ក្តាមដែលមានថ្ងាសតូចចង្អៀតនិងធំទូលាយត្រូវបានដាក់នៅក្នុងអាងចិញ្ចឹមត្រី។ ទឹកមានសារធាតុផ្សំនៃដីឥដ្ឋដែលនៅមានសភាពរំជើបរំជួលដោយមានជំនួយពីម៉ាស៊ីនកូរ។ ក្តាមសរុបចំនួន ២៤៨ ក្បាលត្រូវបានដាក់ក្នុងអាងចិញ្ចឹមត្រី។ មិនយូរប៉ុន្មាន ក្តាមមួយចំនួន (154) បានស្លាប់ ហើយវាបានប្រែក្លាយថាពួកវាទាំងអស់ជារបស់ក្រុម "ចិត្តទូលាយ" ខណៈពេលដែលអ្នកនៅរស់រានមានជីវិត 94 នាក់ទៀតជាក្រុម "ចិត្តចង្អៀត" ។ វាត្រូវបានគេរកឃើញថានៅពេលក្រោយ ការចម្រោះទឹកនៅក្នុងប្រហោងធ្មេញគឺល្អឥតខ្ចោះជាងនៅក្នុង "មុខធំទូលាយ" ដែលជាហេតុផលសម្រាប់ការស្លាប់របស់អ្នកក្រោយ។ ដូច្នេះ នៅក្រោមលក្ខខណ្ឌនៃបាតស្អាត ទម្រង់ "ចិត្តចង្អៀត" មិនមានអត្ថប្រយោជន៍ទេ ហើយសមាមាត្របរិមាណមិនពេញចិត្តទេ។ នៅពេលដែលលក្ខខណ្ឌបានផ្លាស់ប្តូរ ការជ្រើសរើសសម្រាប់ "ចិត្តចង្អៀត" បានចាប់ផ្តើម។
ឧទាហរណ៍ដែលបានពិពណ៌នាក៏បញ្ចេញពន្លឺលើការលុបបំបាត់ចាប (1) ។ អ្នកនិពន្ធខ្លះចាត់ទុកលទ្ធផលនៃការសង្កេតរបស់ Bampes ជាភស្តុតាងដែលថាការជ្រើសរើសមិនបង្កើតអ្វីថ្មីទេ ប៉ុន្តែគ្រាន់តែរក្សានូវបទដ្ឋានជាមធ្យមប៉ុណ្ណោះ (Berg, 1921)។ ការសង្កេតរបស់ Weldon ផ្ទុយពីនេះ។ ជាក់ស្តែង នៅក្រោមលក្ខខណ្ឌធម្មតានៃតំបន់ដែលបានផ្តល់ឱ្យ បទដ្ឋានជាមធ្យមនៅរស់រានមានជីវិត។ នៅក្រោមលក្ខខណ្ឌផ្សេងទៀត បទដ្ឋានជាមធ្យមអាចត្រូវបានលុបចោល ហើយទម្រង់ដែលបង្វែរនឹងនៅរស់។ វាច្បាស់ណាស់ថាក្នុងអំឡុងពេលនៃភូមិសាស្ត្រនៅពេលដែលលក្ខខណ្ឌផ្លាស់ប្តូរ, ជាក្បួន, ក្រោយមកទៀតនឹងកើតឡើង។ នៅក្រោមលក្ខខណ្ឌថ្មី មុខងារថ្មីនឹងលេចចេញមក។
ការពឹងផ្អែកនៃការវិវត្តន៍លើលក្ខខណ្ឌបរិស្ថានគឺអាចមើលឃើញយ៉ាងច្បាស់ពីឧទាហរណ៍ខាងក្រោម។
3. Harrison (1920) បានសង្កេតមើលសមាមាត្រជាលេខនៃបុគ្គលនៃមេអំបៅ Oporabia autumnata ដែលរស់នៅក្នុងតំបន់ព្រៃឈើពីរផ្សេងគ្នានៅក្នុងតំបន់ Cleveland (Yorkshire ប្រទេសអង់គ្លេស)។ យោងតាមលោក Harrison ប្រហែលឆ្នាំ 1800 ។ ព្រៃចម្រុះមានស្រល់, birch និង alder ត្រូវបានបែងចែកជាពីរផ្នែក។ បន្ទាប់ពីខ្យល់ព្យុះនៅពាក់កណ្តាលព្រៃភាគខាងត្បូង ស្រល់ខ្លះបានស្លាប់ ហើយពួកគេត្រូវបានជំនួសដោយដើមប៊ីច។ ផ្ទុយទៅវិញ នៅភាគខាងជើង ដើមឈើ birch និង alder បានក្លាយជាកម្រ។ ដូច្នេះ ព្រៃបានបែកចេញជាពីរស្ថានីយគឺ ដើមស្រល់គ្របដណ្ដប់នៅមួយ ហើយដើមប៊ីចត្រួតត្រានៅកន្លែងផ្សេងទៀត។
វាស្ថិតនៅក្នុងព្រៃនេះដែលមេអំបៅដែលបានរៀបរាប់បានរស់នៅ។ នៅឆ្នាំ 1907 វាត្រូវបានគេកត់សម្គាល់ឃើញថាចំនួនប្រជាជនរបស់វាត្រូវបានបែងចែកជាពីរទម្រង់ - ស្លាបងងឹតនិងស្លាបស្រាល។ ទីមួយគ្របដណ្តប់នៅក្នុងព្រៃស្រល់ (96%) និងទីពីរ - នៅក្នុងព្រៃ birch (85%) ។ សត្វស្លាប Crepuscular (ពាងពេលយប់) និងប្រចៀវបានស៊ីសត្វល្អិតទាំងនេះ ហើយ Harrison បានរកឃើញស្លាបរបស់មេអំបៅដែលត្រូវបានបំផ្លាញនៅលើកម្រាលឈើ។ វាបានប្រែក្លាយថានៅក្នុងព្រៃស្រល់ងងឹត ស្លាបនៅលើដីជាកម្មសិទ្ធិភាគច្រើននៃទម្រង់ពន្លឺ ទោះបីជាសមាមាត្រលេខនៃពូជងងឹតទៅនឹងពន្លឺនៅក្នុងព្រៃស្រល់គឺ 24:1 ក៏ដោយ។ ដូច្នេះហើយ នៅក្នុងព្រៃងងឹតមួយ សត្វស្លាប និងប្រចៀវបានចាប់យកប្រភេទពន្លឺ ដូចដែលវាកាន់តែគួរឱ្យកត់សម្គាល់។ នៅក្នុងឧទាហរណ៍នេះវាត្រូវបានគេមើលឃើញយ៉ាងច្បាស់ថាការឆ្លើយឆ្លងរវាងពណ៌នៃមេអំបៅនិងពណ៌នៃស្ថានីយ៍របស់វាត្រូវបានរក្សាជានិច្ចដោយសកម្មភាពនៃការជ្រើសរើសធម្មជាតិ។
ឥឡូវនេះ ចូរយើងងាកទៅរកភស្តុតាងពិសោធន៍នៃការជ្រើសរើសធម្មជាតិ។ ក្រោយមកទៀតទាក់ទងនឹងផលប៉ះពាល់ជាចម្បងនៃការការពារនៃពណ៌គ្រីប សេម៉ាទិក និង aposematic និងការធ្វើត្រាប់តាម។
4. Poulton (1899) ពិសោធន៍ជាមួយ urticaria pupae 600 ។ កូនឆ្កែត្រូវបានដាក់នៅលើផ្ទៃខាងក្រោយពណ៌ផ្សេងៗ ទាំងផ្គូផ្គង ឬមិនផ្គូផ្គងពណ៌របស់វា។ វាបានប្រែក្លាយថាប្រសិនបើពណ៌របស់ pupae ផ្គូផ្គងនឹងពណ៌នៃផ្ទៃខាងក្រោយនោះចំនួនសរុប 57% ត្រូវបានបំផ្លាញដោយសត្វស្លាបខណៈពេលដែលនៅលើផ្ទៃខាងក្រោយមិនសមរម្យប្រឆាំងនឹង pupae អាចមើលឃើញយ៉ាងច្បាស់ 90% ត្រូវបានបំផ្លាញ។ ការពិសោធន៍ស្រដៀងគ្នានេះត្រូវបានធ្វើឡើងដោយ Cesnola (di-Cesnola, 1904) ដែលបង្ហាញថា mantises ដាក់នៅលើផ្ទៃខាងក្រោយដែលមិនស៊ីគ្នានឹងពណ៌របស់វាត្រូវបានបំផ្លាញទាំងស្រុងដោយសត្វស្លាប។ ទោះយ៉ាងណាក៏ដោយ បច្ចេកទេសរបស់អ្នកស្រាវជ្រាវទាំងនេះគឺបឋម។ Cesnola បានពិសោធន៍ជាមួយ mantises អធិស្ឋានមួយចំនួនតូច។
ទិន្នន័យរបស់ Belyaev និង Geller មានភាពជឿជាក់ជាង។
5. Belyaev (1927) ដូច Chesnola បានពិសោធន៍ជាមួយ mantises អធិស្ឋាន។ ផ្ទៃដីទំហំ ១២០ ម ២ ត្រូវបានបោសសំអាត រុក្ខជាតិខ្ពស់។និងទទួលបានពណ៌ត្នោតរសាត់។ សត្វកន្ធាយចំនួន 60 ក្បាលត្រូវបានដាក់នៅលើទីតាំង ដោយចងជាប់នឹងបង្គោលដែលរុញចូលទៅក្នុងដីនៅចម្ងាយ 1 ម៉ែត្រពីគ្នាទៅវិញទៅមក។ mantises មានពណ៌ត្នោត ចំបើង-លឿង និងបៃតង ហើយ mantises ពណ៌ត្នោតពិបាកនឹងមើលឃើញប្រឆាំងនឹងផ្ទៃខាងក្រោយពណ៌ត្នោតរសាត់នៃគេហទំព័រ។ អ្នកប្រយុទ្ធជាស្រូវសាលី ដែលស្នាក់នៅលើរបងកន្លែងនោះ ហើយបានស៊ីសត្វកន្ធាយ។ ដូច្នេះការពិសោធន៍បង្ហាញយ៉ាងច្បាស់អំពីដំណើរការជ្រើសរើស។
ទិន្នន័យស្រដៀងគ្នានៅលើសម្ភារៈដ៏ធំមួយត្រូវបានបង្ហាញដោយ Heller (1928) ។ សត្វល្អិតត្រូវបានគេដាំនៅលើដីពិសោធតាមលំនាំក្តារខៀន។ អ្នកកំចាត់គឺមាន់។
ការជ្រើសរើសច្បាស់លាស់មួយបានកើតឡើង ដោយសារសត្វល្អិតដែលមិនត្រូវនឹងពណ៌របស់ដីត្រូវបានបំផ្លាញ 95.2% ហើយក្នុងករណី homochromia ផ្ទុយទៅវិញ 55.8% បានរស់រានមានជីវិត។
ការពិសោធន៍របស់ Belyaev និង Geller គឺគួរឱ្យចាប់អារម្មណ៍ក្នុងន័យមួយផ្សេងទៀត: ពួកគេបង្ហាញថា homochromy មិនផ្តល់នូវការធានាពេញលេញនៃការរស់រានមានជីវិតនោះទេប៉ុន្តែគ្រាន់តែបង្កើនសក្តានុពល biotic នៃទម្រង់ដែលបានផ្តល់ឱ្យ។ ជាចុងក្រោយ មានចំណុចមួយទៀតដែលត្រូវគូសបញ្ជាក់។ Mantises ជាកម្មសិទ្ធិរបស់ប្រភេទសត្វដូចគ្នា ហើយភាពខុសគ្នានៃពណ៌របស់ពួកវាគឺមានភាពខុសប្លែកគ្នាក្នុងភាពជាក់លាក់។ ការពិសោធន៍របស់ Belyaev និង Geller បានបង្ហាញថាការជ្រើសរើសកើតឡើងនៅក្នុងចំនួនប្រជាជននៃប្រភេទមួយ។
6. Carrik (1936) បានពិសោធជាមួយ caterpillars ដោយសង្កេតមើលតម្លៃការពារនៃពណ៌គ្រីប។ ជាឧទាហរណ៍ គាត់បានរកឃើញថា wren មិនបានកត់សម្គាល់ឃើញដង្កូវនាងដែលមានពណ៌គ្រីបទេ។ ទោះយ៉ាងណាក៏ដោយ វាគ្រប់គ្រាន់សម្រាប់ដង្កូវនាង ហើយសត្វក្រៀលបានវាយប្រហារវាភ្លាមៗ។ ការសង្កេតស្រដៀងគ្នានេះត្រូវបានធ្វើឡើងដោយអ្នកនិពន្ធផ្សេងទៀត ហើយពួកគេបង្ហាញថា ការដាក់ពណ៌គ្រីបគឺទាក់ទងយ៉ាងជិតស្និទ្ធទៅនឹងឥរិយាបថគ្រីប (ឥរិយាបថសម្រាក) និងចលនាការពារ។
7. ឧទាហរណ៍ខាងលើបង្ហាញពីអត្ថន័យពិតនៃពណ៌គ្រីប។ ឥឡូវនេះ ចូរយើងបន្តទៅអត្ថន័យនៃការធ្វើត្រាប់តាម។ Mostler (1935) បានព្យាយាមបង្កើតថាតើពណ៌ aposematic និង pseudoaposematic មានឥទ្ធិពលកម្រិតណា។ Mostler បានពិសោធន៍ជាមួយ wasps, bumblebees និង bees ក៏ដូចជារុយដែលធ្វើត្រាប់តាមអតីត។ បរិមាណដ៏ច្រើននៃសម្ភារៈបានបង្ហាញថាសត្វស្លាបជាក្បួនមិនបរិភោគ Hymenoptera លើកលែងតែសត្វស្លាបដែលត្រូវបានសម្របខ្លួនជាពិសេសដែលជាក់ស្តែងត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការឆ្លុះបញ្ចាំងរសជាតិ។ ការឆ្លុះបញ្ចាំងនេះត្រូវបានបង្កើតឡើងជាលទ្ធផលនៃបទពិសោធន៍ផ្ទាល់ខ្លួន។ នៅពេលដែលសត្វស្លាបវ័យក្មេងត្រូវបានគេផ្តល់ឱ្យសត្វរុយដែលធ្វើត្រាប់តាម Hymenoptera ពួកគេបានស៊ីវាដំបូង។ ទោះជាយ៉ាងណាក៏ដោយ នៅពេលដែលពួកគេត្រូវបានផ្តល់ hymenoptera ជាលើកដំបូង ហើយពួកគេបានបង្កើតការឆ្លុះបញ្ចាំងអវិជ្ជមានចំពោះសត្វល្អិតទាំងនេះ ពួកគេបានឈប់ទទួលយកសត្វរុយ។ បទពិសោធន៍បានបង្ហាញពីសារៈសំខាន់នៃពណ៌ aposematic និង pseudoaposematic ដ៏អស្ចារ្យ។
បទពិសោធន៍ខាងក្រោមមានសារៈសំខាន់ជាពិសេស។ Miihlmann (1934) ពិសោធន៍ជាមួយសត្វស្លាប បានប្រើដង្កូវនាងធ្វើជាអាហារ។ ដង្កូវត្រូវបានលាបដោយថ្នាំលាបមិនបង្កគ្រោះថ្នាក់ ហើយសត្វស្លាបបានស៊ីវាយ៉ាងងាយ។ បន្ទាប់ពីនេះ បទពិសោធន៍ត្រូវបានកែប្រែ។ សត្វស្លាបត្រូវបានផ្តល់ជូនដង្កូវពណ៌ដូចគ្នា ប៉ុន្តែពួកវាខ្លះត្រូវបានលាបពណ៌ដោយលាយថ្នាំលាប និងសារធាតុរសជាតិមិនល្អ។ សត្វស្លាបបានឈប់យកដង្កូវបែបនេះ ប៉ុន្តែពួកគេមិនយកពណ៌ធម្មតាទេ ពោលគឺអាចបរិភោគបាន។ ទំនាក់ទំនងមួយបានកើតឡើងដែលមានលក្ខណៈស្រដៀងគ្នារវាងអ្នកត្រាប់តាមនិងគំរូ។ អ្នកដែលលាបពណ៌ដោយល្បាយមិនសប្បាយចិត្តដើរតួជាគំរូ អ្នកដែលលាបយ៉ាងសាមញ្ញគឺជាអ្នកត្រាប់តាម។ ដូច្នេះវាត្រូវបានបង្ហាញថាភាពស្រដៀងគ្នានៃអ្នកត្រាប់តាមគំរូមានតម្លៃការពារ។ បន្ទាប់មកការពិសោធន៍ត្រូវបានកែប្រែដូចខាងក្រោម។ Mühlmann បានកំណត់ដើម្បីស្វែងយល់ថាតើសត្វស្លាបអាចបែងចែកគំរូបានដល់កម្រិតណា។ ថ្នាំលាបត្រូវបានអនុវត្តទៅផ្នែកខ្លះនៃដងខ្លួនរបស់ដង្កូវ ពួកគេត្រូវបានផ្តល់គំរូជាក់លាក់មួយ ហើយក្នុងទម្រង់នេះ ដង្កូវត្រូវបានរួមបញ្ចូលនៅក្នុងការពិសោធន៍ដែលបានពិពណ៌នាខាងលើ។ វាបានប្រែក្លាយថាសត្វស្លាបបានសម្គាល់គំនូរហើយមិនយកដង្កូវលាបពណ៌ទេប្រសិនបើក្រោយមកទៀតមានរសជាតិមិនល្អ។ លទ្ធផលនេះបញ្ចេញពន្លឺលើដំណើរការនៃការកែលម្អគំនូរគ្រីប។ ប្រសិនបើសត្វស្លាបបែងចែកគំរូមួយ នោះកាន់តែល្អឥតខ្ចោះ ឧទាហរណ៍ ភាពស្រដៀងនឹងស្លាបមេអំបៅទៅនឹងស្លឹក នោះឱកាសនៃការរស់រានមានជីវិតកាន់តែច្រើន។ នៅក្នុងពន្លឺនៃការពិសោធន៍របស់Mühlmann ការសន្និដ្ឋាននេះទទួលបានកម្រិតខ្ពស់នៃភាពជឿជាក់។
ការជ្រើសរើសផ្លូវភេទ
ទ្រឹស្ដីនៃការជ្រើសរើសផ្លូវភេទបានបង្កើតការជំទាស់ច្រើនបំផុតសូម្បីតែពី Darwinists ជាច្រើនក៏ដោយ។ វាបានប្រែក្លាយថានៅក្នុងករណីមួយចំនួននៃការប្រើប្រាស់របស់វាអាចមានការជំទាស់ ហើយជាឧទាហរណ៍ ពណ៌ភ្លឺរបស់បុរសអាចត្រូវបានពន្យល់ខុសគ្នា។ ដូច្នេះ Wallace បានសន្មត់ថាពណ៌ និងលំនាំមិនប៉ះពាល់ដល់ជម្រើសរបស់ស្ត្រីទេ ហើយថាកម្លាំងរបស់បុរសគឺមានសារៈសំខាន់បំផុត ដែលត្រូវបានបង្ហាញជាពណ៌ភ្លឺជាង។ ដូច្នេះ Wallace បដិសេធការជ្រើសរើសផ្លូវភេទជាសំខាន់។ ពួកគេបានព្យាយាមបដិសេធទ្រឹស្ដីនៃការជ្រើសរើសផ្លូវភេទដោយហេតុផលថាវាផ្អែកលើមនុស្សនិយម ពោលគឺលើការផ្ទេរមេកានិកនៃអារម្មណ៍របស់មនុស្សទៅសត្វ។ ការបន្ថែមមេកានិកនៃគំនិតរបស់មនុស្សអំពីភាពស្រស់ស្អាតដល់សត្វគឺពិតជាខុសឆ្គង។ ជាការពិតណាស់ យើងមិនដឹងថា ទួរគី "គិត" យ៉ាងណាទេ អំពីទួរគីដែលអួតពីមុខនាង ប៉ុន្តែយើងមិនអាចផ្អែកលើការសង្កេតសាមញ្ញ បដិសេធ ឬការពារទ្រឹស្ដីនៃការជ្រើសរើសផ្លូវភេទនោះទេ។ Zhitkov (1910) ដោយផ្អែកលើការសង្កេតលើវាលជាច្រើន ចង្អុលបង្ហាញថា ការរួមរ័ករបស់ grouse ខ្មៅ និងការប្រយុទ្ធរបស់ turukhtans ជាញឹកញាប់កើតឡើងដោយគ្មានការចូលរួមពីស្ត្រី ហើយដូច្នេះមិនមានជម្រើសសម្រាប់បុរសទេ។ Zhitkov ក៏បានចង្អុលបង្ហាញថានៅក្នុង grouse leks បុរសសកម្មបំផុតប្រយុទ្ធនៅផ្នែកកណ្តាលនៃ lek ។ នៅសល់ ទន់ខ្សោយ និងក្មេងជាង ស្នាក់នៅជាយក្រុងរបស់វា កាន់តែជិតស្រី ដែលជាមូលហេតុ "ជាមួយនឹងកម្រិតនៃប្រូបាប៊ីលីតេកាន់តែច្រើន វាអាចសន្មត់ថាពួកគេតែងតែទទួលបានការចាប់អារម្មណ៍ពីស្ត្រី" ។
ការពិតបែបនេះហាក់ដូចជានិយាយប្រឆាំងនឹងទ្រឹស្តីនៃការជ្រើសរើសផ្លូវភេទ។ វាត្រូវបានគេណែនាំផងដែរថាពណ៌ភ្លឺរបស់បុរសមិនមានតម្លៃគួរឱ្យទាក់ទាញទេប៉ុន្តែគួរឱ្យភ័យខ្លាច។ Fausek (1906) បានបង្កើតទ្រឹស្តីនេះយ៉ាងលម្អិត។ គ្មានការងឿងឆ្ងល់ទេដែលថាទ្រឹស្តីនៃការបំភ័យ (ការគំរាមកំហែង) ការលាបពណ៌មិនអាចបដិសេធបានទេ។
ទោះជាយ៉ាងណាក៏ដោយ វាគួរតែត្រូវបាននិយាយថា ការពិចារណាទាំងនេះមិនបដិសេធទ្រឹស្តីនៃការជ្រើសរើសផ្លូវភេទជាសំខាន់នោះទេ។ នេះទាក់ទងជាចម្បងទៅនឹងការសង្កេតខាងលើរបស់ Zhitkov ដែលយោងទៅតាមការបង្ហាញ grouse ខ្មៅសូម្បីតែនៅក្នុងអវត្តមាននៃស្ត្រីហើយការប្រយុទ្ធជាមួយ grouse ខ្មៅ (grouse ខ្មៅរបស់បុរស) មិនយកចិត្តទុកដាក់ចំពោះស្ត្រីទោះបីជាពួកគេមានវត្តមានក៏ដោយ។ ការសង្កេតដំបូងបង្ហាញតែថា ការសម្របខ្លួនទៅនឹងរដូវកាលមិត្តរួមគឺទាក់ទងទៅនឹងការសម្របខ្លួនណាមួយ។ អាកប្បកិរិយារបស់ត្រីបាឡែនខ្មៅនៅលើ lek ក្លាយជាការសម្របខ្លួននៅក្នុងវត្តមាននៃទំនាក់ទំនងជាក់លាក់មួយពោលគឺនៅក្នុងវត្តមានរបស់ស្ត្រី។ នៅក្នុងទំនាក់ទំនងផ្សេងទៀត បាតុភូតដូចគ្នាមិនមានអត្ថន័យនៃការសម្របខ្លួនទៅនឹងរដូវកាលមិត្តរួមនោះទេ។ ការសង្កេតនេះដោយ Zhitkov មិនបញ្ជាក់ពីអ្វីផ្សេងទៀតទេ។ ទាក់ទងនឹងការសង្កេតលើកទីពីររបស់គាត់ឥឡូវយើងដឹងច្បាស់ហើយ។ ឥទ្ធិពលផ្ទាល់បង្ហាញពីភាពស្រើបស្រាលផ្លូវភេទចំពោះបុរស និងស្ត្រី. មនុស្សម្នាក់អាចគិតថា វាគឺជាបុរសដែលបង្ហាញភាពស្រើបស្រាល បន្ទាប់ពីបង្ហាញរួច ស្ថិតក្នុងស្ថានភាពនៃការកើនឡើងនូវភាពស្រើបស្រាលនៃការរួមភេទ ដែលកាន់តែខិតជិតមនុស្សស្រីយ៉ាងសកម្ម ហើយវាគឺជាពួកគេដែលទទួលបានជោគជ័យខ្លាំងបំផុត ខណៈដែលបុរសដែលមិនចូលរួមក្នុងការបង្ហាញ និងប្រយុទ្ធ ដោយសារតែ ដល់ការខ្វះការស្រើបស្រាលខាងផ្លូវភេទ ចូរនៅខាងក្រៅ។ ដូច្នេះក្នុងករណីនៃ grouse ខ្មៅ យើងប្រហែលជាកំពុងដោះស្រាយជាមួយនឹងទម្រង់នៃការជ្រើសរើសផ្លូវភេទ ដែលបុរសជាភាគីសកម្ម។ ទម្រង់នៃការជ្រើសរើសផ្លូវភេទនេះគឺពិតជាករណីពិសេសនៃការជ្រើសរើសតាមធម្មជាតិ។ កម្លាំងរបស់បុរស អាវុធរបស់គាត់ ការសម្របខ្លួនរបស់គាត់សម្រាប់ការការពារ និងការវាយប្រហារសកម្មគឺមានសារៈសំខាន់យ៉ាងធំធេងក្នុងការតស៊ូដើម្បីអត្ថិភាព។ ជាឧទាហរណ៍ ចង្កូមធំអាចមានសារៈសំខាន់ទាំងក្នុងការប្រយុទ្ធស្រី និងការពារសត្រូវ។ ដូច្នេះ ក្នុងករណីបែបនេះ យើងអាចនិយាយអំពីភាពចៃដន្យនៃការជ្រើសរើសផ្លូវភេទ និងធម្មជាតិ ហើយការរួមរស់ជាមួយបុរសដែលស្វាហាប់ និងរឹងមាំជាងមុន (ប្រសិនបើលក្ខណៈ និងលក្ខណៈសម្បត្តិរបស់គាត់ត្រូវបានកំណត់តាមតំណពូជ) ជាការពិតណាស់ ការកើនឡើង ស្តង់ដារនៃការរស់នៅ ចំនួនប្រជាជនដែលកើតចេញពីបុរសបែបនេះ។ យើងពិតជាសង្កេតឃើញទម្រង់នៃការជ្រើសរើសផ្លូវភេទនេះនៅក្នុងថនិកសត្វដែលមានការរៀបចំខ្ពស់ ( canines , deer , seals ) និងនៅក្នុងបក្សី។ ប្រសិនបើក្នុងករណីនេះបាតុភូតដែលបានពិពណ៌នាដោយ Zhitkov កើតឡើងនោះមនុស្សម្នាក់មិនអាចបំភ្លេចទំនាក់ទំនងនៃការសម្របខ្លួនណាមួយបានទេហើយរំពឹងថាកម្លាំងកាន់តែច្រើននិងអាវុធកាន់តែប្រសើរឡើងក្នុងគ្រប់ករណីទាំងអស់ធានាបាននូវមិត្តរួមរបស់បុរសពិសេសទាំងនេះ មិនមែនរបស់ផ្សេងទៀតដែលខ្សោយជាងនោះទេ។ ទីពីរ នៅពេលពិភាក្សាអំពីការពិតនៃទម្រង់នៃការជ្រើសរើសផ្លូវភេទនៅក្នុងសំណួរ កត្តាមួយបន្ថែមទៀតត្រូវតែយកមកពិចារណាគឺ កម្ពស់របស់អង្គការ។ ជាឧទាហរណ៍ វាមិនអាចទៅរួចទេក្នុងការ "បដិសេធ" ទ្រឹស្ដីនៃការជ្រើសរើសផ្លូវភេទ ដោយប្រើឧទាហរណ៍ពីទំនាក់ទំនងរវាងភេទក្នុងទម្រង់រៀបចំទាប។ និយាយយ៉ាងតឹងរ៉ឹង ការជ្រើសរើសផ្លូវភេទ ផ្ទុយទៅនឹងការជ្រើសរើសធម្មជាតិ កើតឡើងតាមរយៈការជ្រើសរើសបុគ្គលដែលសមស្រប ហើយដូច្នេះវាត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការអភិវឌ្ឍខ្ពស់នៃប្រព័ន្ធសរសៃប្រសាទ និងសរីរាង្គអារម្មណ៍។ ដូច្នេះ គេអាចប្រកែកបានថា សារៈសំខាន់នៃការជ្រើសរើសផ្លូវភេទកើនឡើងនៅពេលដែលអង្គការកើនឡើង។ តាមទស្សនៈនេះ ការប៉ុនប៉ងរបស់ J. S. Huxley (1940) ដើម្បីចូលទៅជិតទំនាក់ទំនងរវាងភេទតាមទស្សនៈប្រវត្តិសាស្ត្រគឺគួរឱ្យចាប់អារម្មណ៍។ គាត់បែងចែកក្រុមសំខាន់ៗចំនួនបីដូចខាងក្រោមនៃទំនាក់ទំនងទាំងនេះ។ ក - ទម្រង់ដោយគ្មានការឆ្លងកាត់ ដែលក្នុងនោះ gametes ត្រូវបានរួបរួមដោយមិនគិតពីទំនាក់ទំនងណាមួយរវាងបុគ្គល ឧទាហរណ៍ដោយការបញ្ចេញពង និងមេជីវិតឈ្មោលទៅក្នុងទឹក ដូចដែលយើងឃើញនៅក្នុង coelenterates, annelids និងត្រី teleost ភាគច្រើន។ តាមធម្មជាតិ មិនអាចមានការនិយាយអំពីការជ្រើសរើសផ្លូវភេទនៅទីនេះទេ។ ខ - ទម្រង់ជាមួយមិត្តរួមទោះជាយ៉ាងណាសម្រាប់តែ coitus ដោយគ្មានការរួមរស់យូរអង្វែងនៃការរួមភេទ។ ក្នុងករណីនេះយើងឃើញការអភិវឌ្ឍឧបករណ៍ពិសេសដែលទាក់ទាញភេទទាំងពីរឱ្យគ្នាទៅវិញទៅមក។ នេះរាប់បញ្ចូលទាំងបាតុភូតពីរប្រភេទ៖ ក) ការអភិវឌ្ឍន៍សមត្ថភាពក្នុងការរួមរស់ជាមួយបុគ្គលម្នាក់។ ឧទាហរណ៍៖ ការរកឃើញភេទផ្ទុយដោយប្រើសរីរាង្គនៃក្លិន ការមើលឃើញ ការស្តាប់ ការរំញោចនៃប្រតិកម្មផ្លូវភេទដោយការប៉ះ ឬចាប់ (នៅក្នុងក្តាមខ្លះ ក្នុងសត្វពាហនៈគ្មានកន្ទុយ) ល្បែងផ្លូវភេទដែលជំរុញឱ្យមានមិត្តរួម (newts, dipterans ខ្លះ ។ល។) , ចំបាប់ និងការបំភិតបំភ័យ (ដង្កូវនាង, ជីងចក់, ដំបងគ្រញូង, មេទីសបន់ស្រន់។ល។)។ ខ) ការអភិវឌ្ឍន៍សមត្ថភាពក្នុងការរួមរស់ជាមួយបុគ្គលច្រើនជាងមួយរូប ដោយមានជំនួយពី៖ ក) ចំបាប់ ខ) មិត្តរួម គ) ចំបាប់ និងមិត្តរួម (ដូចត្រូវបានគេសង្កេតឃើញនៅក្នុង ក្ងោកក្រហម ក្អែកខ្មៅ បក្សីឋានសួគ៌)។ គ - ការរួមរស់បានយូរនៃការរួមភេទមិនត្រឹមតែក្នុងអំឡុងពេល coitus ប៉ុណ្ណោះទេប៉ុន្តែថែមទាំងក្នុងអំឡុងពេលទំនាក់ទំនងបន្ថែមទៀតផងដែរ។ ការរួមភេទកើតឡើង៖ ក) ជាមួយបុគ្គលម្នាក់ ឬ ខ) ជាមួយបុគ្គលជាច្រើន ហើយការរួមរស់ត្រូវបានភ្ជាប់ជាមួយនឹងការប្រយុទ្ធ ឬការប្រយុទ្ធរួមគ្នាជាមួយនឹងការទាក់ទាញការចាប់អារម្មណ៍ ។ល។ e. នេះរួមបញ្ចូលទាំងទំនាក់ទំនងរវាងភេទនៅក្នុងថ្នាក់នៃសត្វស្លាប និងថនិកសត្វ។
គ្រោងការណ៍របស់ Huxley គឺផ្អែកលើការវិវឌ្ឍន៍នៃប្រព័ន្ធសរីរាង្គបន្តពូជ ហើយនេះគឺជាគុណវិបត្តិរបស់វា។ វានឹងជាការត្រឹមត្រូវជាងក្នុងការកសាងគ្រោងការណ៍នេះលើការអភិវឌ្ឍន៍ជាលំដាប់នៃប្រព័ន្ធសរសៃប្រសាទ។ តាមការពិត វាមិនត្រឹមត្រូវទេក្នុងការដាក់ហ្គេមផ្លូវភេទលើសត្វរុយ និងរុយផ្លែឈើ និងទំនាក់ទំនងរវាងសត្វកន្លាតឈ្មោល និងសត្វចៃក្រោមចំណងជើងដូចគ្នា។ ប្រសិនបើយើងចាត់ថ្នាក់ទំនាក់ទំនងរវាងភេទដោយយោងទៅតាមកម្រិតនៃការអភិវឌ្ឍន៍ប្រព័ន្ធសរសៃប្រសាទ យើងអាចបញ្ជាក់ថា ការជ្រើសរើសផ្លូវភេទ ក្នុងទម្រង់ធម្មតារបស់វា បង្ហាញរាងដោយខ្លួនវាផ្ទាល់នៅក្នុងសត្វខ្ពស់ជាង (សត្វឆ្អឹងខ្នង ជាពិសេសសត្វស្លាប និងថនិកសត្វ) ដែលមានសមត្ថភាពនៃសកម្មភាពឆ្លុះបញ្ចាំងតាមលក្ខខណ្ឌ។
យើងគ្រាន់តែត្រូវចងចាំទំនាក់ទំនងនៃការជ្រើសរើសផ្លូវភេទ។ ជាឧទាហរណ៍ បុរសខ្លាំងបំផុតនឹងមិនតែងតែជោគជ័យខ្លាំងបំផុតនោះទេ។ នៅ grouse leks, coitus មិនតែងតែត្រូវបានផ្តល់ឱ្យសម្រាប់តែបុរសដែលចូលរួមក្នុងការមិត្តរួមនោះទេ។ ប៉ុន្តែជាមធ្យម បុរសដែលខ្លាំងបំផុត និងសកម្មបំផុតនៅតែមានឱកាសល្អជាងបុរសដែលនៅសល់។ ការរិះគន់លើទ្រឹស្ដីនៃការជ្រើសរើសប្រភេទទីមួយ ដែលការរួមរស់អាស្រ័យទៅលើការប្រកួតប្រជែងរវាងបុរស គឺផ្អែកលើការបកស្រាយខុសនៃទ្រឹស្តីនៃការសម្របខ្លួន។ អ្នករិះគន់ដាក់លើ Darwinism នូវគំនិតនៃសារៈសំខាន់ដាច់ខាតនៃការសម្របខ្លួន ហើយបន្ទាប់មកដោយការលើកឡើងពីករណីដែលការសម្របខ្លួនបែបនេះមិនមានសុពលភាព អះអាងថាពួកគេគ្មានន័យអ្វីទាំងអស់។ តាមពិត ការសម្របខ្លួនណាមួយ ដូចដែលយើងដឹងគឺទាក់ទងគ្នា ហើយដូច្នេះការជ្រើសរើសផ្លូវភេទមិនតែងតែធ្វើតាមគ្រោងការណ៍ដែលស្នើឡើងដោយដាវីននោះទេ។
ចំណុចកណ្តាលនៃទ្រឹស្តីនៃការជ្រើសរើសផ្លូវភេទគឺជាបញ្ហានៃពណ៌ភ្លឺរបស់ឈ្មោលនៃសត្វស្លាបជាច្រើន (និងសត្វដទៃទៀត ប៉ុន្តែជាពិសេសសត្វស្លាប)។ យ៉ាងណាមិញ វាគឺជាពណ៌ដ៏ភ្លឺស្វាងរបស់បុរស ដែលផ្ទុយនឹងទ្រឹស្តីនៃការជ្រើសរើសធម្មជាតិ ដែលទាមទារការពន្យល់។ Darwin បានដាក់ចេញនូវទ្រឹស្ដីដ៏ប៉ិនប្រសប់មួយ យោងទៅតាមស្ត្រីជ្រើសរើសបុរសដែលស្អាតជាងគេ។ ទ្រឹស្តីនេះអាចត្រូវបានបដិសេធ ឬបញ្ជាក់ដោយពិសោធន៍តែប៉ុណ្ណោះ។ មានទិន្នន័យតិចតួចអំពីបញ្ហានេះ។ ទោះជាយ៉ាងណាក៏ដោយ យើងបង្ហាញលទ្ធផលខាងក្រោមនៃការសង្កេតពិសោធន៍ (Cinat - Thomson, 1926) លើការជ្រើសរើសផ្លូវភេទនៅក្នុង budgerigar (Melopsittacus undulatus)។ ឈ្មោលនៃបក្សីនេះមានរោមខៀវស្រងាត់បង្កើតជាកអាវដែលមានចំណុចងងឹតធំ ៗ (1-5) ឬ 1-3 តូចជាង។ ចំណុចកាន់តែច្រើន កអាវកាន់តែអភិវឌ្ឍកាន់តែប្រសើរ។ បើតាមចំនួនចំណុចបុរសត្រូវបានកំណត់លេខ១ លេខ២ លេខ៣ ។ល។ ឈ្មោលលេខ 2 និងលេខ 4 ត្រូវបានដាក់ក្នុងទ្រុង ស្ត្រីទាំងអស់ជ្រើសរើសឈ្មោលលេខ 4 ។ បន្ទាប់មកការពិសោធន៍ខាងក្រោមត្រូវបានធ្វើ។ សត្វឈ្មោលមានរោមខ្មៅបន្ថែមទៀតដែលជាប់នឹងកអាវរបស់ពួកគេ។ ឈ្មោលលេខ 4 លេខ 3 លេខ 2 និងលេខ 1 ត្រូវបានទទួលរងនូវការពិសោធន៍ ការពិសោធន៍ត្រួតពិនិត្យបានបង្ហាញថាស្ត្រីជ្រើសរើសឈ្មោលលេខ 3 និងលេខ 4 ។ បុរសទាំងនេះត្រូវបានទុកចោលនៅក្នុង plumage ធម្មជាតិរបស់ពួកគេ។ បន្ទាប់មក "បុរសលាបពណ៌" លេខ 2+1 និងលេខ I + II (លេខរ៉ូម៉ាំងបង្ហាញពីចំនួនរោមដែលស្អិតជាប់) ត្រូវបានបញ្ចេញទៅក្នុងឯករភជប់។ ទោះបីជាជោគជ័យរបស់ពួកគេទាបជាងការរំពឹងទុកក៏ដោយ ក៏វានៅតែក្លាយជាជោគជ័យទ្វេដងរបស់ពួកគេ (នៅពេលដែលបុរសទាំងនេះមិនមានរោមសត្វស្អិតជាប់)។ នៅក្នុងការពិសោធន៍មួយផ្សេងទៀត បុរសលេខ 4 (ដែលទទួលបានជោគជ័យ) បានកាត់កអាវរោមរបស់គាត់ចេញ ហើយរោមខ្មៅនៅលើវាក៏ដកចេញ។ គាត់ត្រូវបានអនុញ្ញាតឱ្យចូលទៅក្នុងឯករភជប់ ហើយជាការបរាជ័យទាំងស្រុង។ ទោះបីជាមានភាពមិនត្រឹមត្រូវនៃវិធីសាស្រ្ត (ទិន្នន័យនឹងមានភាពត្រឹមត្រូវជាងមុនដោយប្រើស្ថិតិបំរែបំរួល) យ៉ាងណាក៏ដោយ ការពិសោធន៍ទាំងនេះបង្ហាញថាស្ត្រីបែងចែក និងជ្រើសរើសបុរសដោយផ្អែកលើរូបរាងរបស់ពួកគេ។
ដូច្នេះអត្ថិភាពនៃការជ្រើសរើសផ្លូវភេទត្រូវបានបង្កើតឡើងដោយពិសោធន៍។ វាគួរតែត្រូវបានសង្កត់ធ្ងន់ថានៅក្នុងការពិសោធន៍របស់ Cinat-Thomson ស្ត្រីជ្រើសរើសបុរសដែលបញ្ជាក់ពីទីតាំងកណ្តាលនៃទ្រឹស្តីនៃការជ្រើសរើសផ្លូវភេទជាកត្តាកំណត់ពណ៌ភ្លឺរបស់បុរស។
បញ្ហានៃការជ្រើសរើសផ្លូវភេទថ្មីៗនេះបានទទួលការគ្របដណ្តប់គួរឱ្យចាប់អារម្មណ៍នៅក្នុងស្នាដៃរបស់អ្នកនិពន្ធមួយចំនួនរួមទាំង Mashkovtsev ដែលផ្អែកលើទិន្នន័យអក្សរសាស្ត្រនិងការសង្កេតផ្ទាល់ខ្លួនរបស់គាត់ (Mashkovtsev, 1940) បានឈានដល់ការសន្និដ្ឋានថាវត្តមានរបស់បុរសមាន ឥទ្ធិពលរំញោចលើការវិវត្តនៃអូវែរ និងចំនួនពងក្នុងស្ត្រី បរិស្ថានទូទៅក៏មានសារៈសំខាន់ផងដែរ។ រដូវកាលមិត្តរួមវត្តមាននៃសំបុក រូបរាងនៃរុក្ខជាតិបៃតងនិទាឃរដូវ បំណះដែលរលាយ។ ផ្ទុយទៅវិញ ប្រសិនបើអ្នកបង្កើតសំបុក ហើយឱ្យឈ្មោលចូល នោះការបញ្ចេញពងអូវុលយ៉ាងឆាប់រហ័ស (ការវិវត្តនៃពង) និងការវិវត្តន៍ដែលពឹងផ្អែកខ្លាំងនៃអូវែរចាប់ផ្តើម។ ដូច្នេះកត្តាបរិស្ថានខាងក្រៅក៏ដូចជាសំបុកនិងបុរស (ក្លិននិងរូបរាងរបស់គាត់) មានឥទ្ធិពលលើស្ត្រីរំញោច oogenesis ។ ប្រសិនបើយើងប្រៀបធៀបទិន្នន័យទាំងនេះយ៉ាងហោចណាស់ជាមួយនឹងការពិសោធន៍របស់ Cinat Thomson វាច្បាស់ណាស់ថាសរីរាង្គនៃអារម្មណ៍ (ជាចម្បងសរីរាង្គនៃចក្ខុវិស័យ) នៅក្នុងសត្វស្លាបមានសារៈសំខាន់យ៉ាងខ្លាំងក្នុងការកើតឡើងនៃអារម្មណ៍ផ្លូវភេទចំពោះស្ត្រី។ សញ្ញារបស់បុរស (ក៏ដូចជាវត្តមាននៃសំបុក និងស្ថានភាពអេកូឡូស៊ីដែលត្រូវគ្នា) តាមរយៈអារម្មណ៍ ជាក់ស្តែងជំរុញសកម្មភាពនៃក្រពេញភីតូរីសរបស់ស្ត្រី ដែលបញ្ចេញអរម៉ូន gonadotropic (រំញោចមុខងារអូវែ) ។ យើងឃើញថាការភ្ញោចពីខាងក្រៅ និងជាពិសេសវត្តមានរបស់បុរស គឺជាកត្តាដ៏មានឥទ្ធិពលដែលជួយបង្កើនការផលិតផ្លូវភេទរបស់ស្ត្រី។ ទិន្នន័យដែលបានបង្ហាញពិតជាបញ្ជាក់ពីបទប្បញ្ញត្តិសំខាន់ៗនៃទ្រឹស្ដីការជ្រើសរើសផ្លូវភេទរបស់ដាវីន។ ក្នុងករណីនេះ វាមានលទ្ធភាពខ្ពស់ដែលការជ្រើសរើសផ្លូវភេទ ដែលជាទម្រង់ពិសេសនៃការជ្រើសរើសធម្មជាតិ ដើរតួនាទីយ៉ាងធំជាកត្តាបង្កើនការមានកូនរបស់ស្ត្រី។ ការកើនឡើងនៃអត្រាបន្តពូជ (ក្រោមលក្ខខណ្ឌទូទៅអំណោយផលមួយចំនួន) នាំឱ្យមានការកើនឡើងនៃសក្តានុពលជីវសាស្ត្រទូទៅនៃប្រភេទសត្វ។ ទំនាក់ទំនងទាំងនេះដកចេញនូវសារសំខាន់អវិជ្ជមាននៃពណ៌ដែលមិនលាក់បាំងរបស់បុរស ហើយវាក្លាយជាកត្តាមួយក្នុងការអភិវឌ្ឍន៍រីកចម្រើន និងជោគជ័យក្នុងជីវិតរបស់ប្រភេទសត្វ។
ការជ្រើសរើសផ្លូវភេទ និងភាពខុសគ្នាខាងផ្លូវភេទ. ពីបទបង្ហាញមុនវាច្បាស់ណាស់ថាការជ្រើសរើសផ្លូវភេទត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងភាពខុសគ្នា morphophysiological រវាងបុរសនិងស្ត្រី។ វាត្រូវបានគេដឹងថាបុរស និងស្ត្រីមានភាពខុសប្លែកគ្នានៅក្នុងលក្ខណៈផ្លូវភេទបន្ទាប់បន្សំរបស់ពួកគេ ហើយក្រោយមកទៀតទាំងនេះកើតឡើងក្រោមឥទ្ធិពលនៃអរម៉ូនភេទបុរស និងស្ត្រីដែលផលិតនៅក្នុង gonads ។ ការពិសោធន៍ជាមួយនឹងការប្តូរ gonads ពីបុរសទៅស្ត្រី និងពីក្រោយទៅបុរស បង្ហាញឱ្យឃើញពីការពឹងផ្អែកនៃលក្ខណៈផ្លូវភេទបន្ទាប់បន្សំលើសកម្មភាពអ័រម៉ូនរបស់ gonads ។ ទំនាក់ទំនងទាំងនេះហាក់ដូចជាធ្វើឱ្យវាអាចធ្វើទៅបានដើម្បីកាត់បន្ថយភាពចម្រុះផ្លូវភេទទៅនឹងឥទ្ធិពលអ័រម៉ូនសុទ្ធសាធ និងមើលឃើញពីហេតុផលនៃភាពខុសគ្នារវាងបុរស និងស្ត្រី។ ជាមួយនឹងការបង្កើតសំណួរនេះ ទ្រឹស្តីនៃការជ្រើសរើសផ្លូវភេទហាក់ដូចជាមិនចាំបាច់។ ជាការពិតណាស់នៅដំណាក់កាលទាបនៃការអភិវឌ្ឍន៍ phylogenetic បញ្ហានៃ dimorphism ផ្លូវភេទអាចត្រូវបានដោះស្រាយដោយផ្អែកលើទ្រឹស្តីនៃឥទ្ធិពលអ័រម៉ូនភេទ។ យើងក៏អាចពិចារណាថា dimorphism ផ្លូវភេទនៅក្នុងករណីទាំងនេះត្រូវបានកំណត់ដោយកត្តាហ្សែន។ ជាឧទាហរណ៍ នៅក្នុងពពួកដង្កូវមូល ភាពចម្រុះផ្លូវភេទត្រូវបានបង្ហាញយ៉ាងច្បាស់ ហើយបុរសអាចបែងចែកយ៉ាងច្បាស់ពីស្ត្រីដោយលក្ខណៈផ្លូវភេទបន្ទាប់បន្សំរបស់ពួកគេ ខណៈពេលដែលវាពិបាកក្នុងការនិយាយអំពីការជ្រើសរើសផ្លូវភេទនៅក្នុងក្រុមនៃសារពាង្គកាយនេះ។ ទាំងការប្រកួតប្រជែងរវាងបុរស និងការជ្រើសរើសបុរសដោយស្ត្រីកើតឡើងនៅទីនេះ ទោះបីជាទំនាក់ទំនងរវាងភេទនៅក្នុង nematodes គួរតែត្រូវបានចាត់ថ្នាក់ក្រោមចំណងជើងទីពីររបស់ J.S. Huxley ក៏ដោយ។ បុរសនិងស្ត្រីចូលទៅក្នុង coitus ដែលមុនដោយបុរសចាប់យករាងកាយរបស់ស្ត្រី។ បុរសរុំកន្ទុយជុំវិញនាង មានអារម្មណ៍ថាប្រដាប់ភេទបើកចេញ ហើយបញ្ចូលសាច់របស់វា បន្ទាប់មកចាក់គ្រាប់ពូជចេញពីប្រឡាយទឹកកាម។ បាតុភូតទាំងនេះមិនត្រូវបានផ្សារភ្ជាប់ជាមួយនឹងការជ្រើសរើសផ្លូវភេទទេ។ ការសង្កេតជាច្រើនរបស់អ្នកនិពន្ធអំពីអាកប្បកិរិយារបស់បុរសបង្ហាញថា coitus កើតឡើងជាលទ្ធផលនៃការជួបនឹងឱកាស។
នៅក្នុងសត្វខ្ពស់ជាង - សត្វឆ្អឹងខ្នង (សត្វល្អិត) និងសូម្បីតែច្រើនទៀតនៅក្នុងសត្វឆ្អឹងខ្នង - ការជ្រើសរើសផ្លូវភេទគឺមិនអាចប្រកែកបាន។ អាស្រ័យហេតុនេះ សំណួរកើតឡើង៖ តើអ្វីជាមូលហេតុនៃភាពខុសប្រក្រតីផ្លូវភេទនៅទីនេះ - ការជ្រើសរើសផ្លូវភេទ ឬឥទ្ធិពលនៃកត្តាអ័រម៉ូន? សំណួរនេះគួរតែត្រូវបានឆ្លើយដូចនេះ។ តាមប្រវត្តិសាស្ត្រ ភាពខុសប្រក្រតីផ្លូវភេទបានកើតឡើងនៅក្នុងទំនាក់ទំនងអ័រម៉ូនរបស់វា។ នោះហើយជាមូលហេតុដែលវាមានវត្តមាននៅក្នុងក្រុមទាបដែលមិនមានការជ្រើសរើសផ្លូវភេទ។ ទោះបីជាយ៉ាងណាក៏ដោយ នៅក្នុងទម្រង់ខ្ពស់ ជាពិសេសនៅក្នុងសត្វស្លាប និងថនិកសត្វ កត្តាអ័រម៉ូនជាប្រវត្តិសាស្ត្រផ្តល់ផ្លូវដល់ការជ្រើសរើសផ្លូវភេទ ហើយភាពខុសប្លែកគ្នាខាងផ្លូវភេទកើតឡើងលើសារៈសំខាន់នៃទម្រង់ពិសេសនៃភាពប្រែប្រួលដែលបម្រើជាសម្ភារៈសម្រាប់ការកើតឡើងនៃការជ្រើសរើសផ្លូវភេទ។ ពណ៌ភ្លឺ កម្លាំង និងអាវុធរបស់បុរសគឺជាផលវិបាកផ្ទាល់នៃឥទ្ធិពលនៃអរម៉ូនភេទ។ ទោះជាយ៉ាងណាក៏ដោយ វាស្ថិតក្រោមឥទ្ធិពលនៃការជ្រើសរើសផ្លូវភេទយ៉ាងជាក់លាក់ ដែលការបន្តពូជបន្តពូជរបស់បុរសទាំងនោះដែលមានសិទ្ធិបន្តពូជ។ លក្ខណៈត្រូវបានអភិវឌ្ឍយ៉ាងពេញលេញ និងច្បាស់លាស់បំផុត។ ដូច្នេះ តាមរយៈការជ្រើសរើសភេទនៃលក្ខណៈខាងក្រៅ ឥទ្ធិពលអ័រម៉ូននៃ gonad ហើយជាលទ្ធផល ការជ្រើសរើសសម្រាប់ dimorphism ផ្លូវភេទកើនឡើង។
ប្រសិនបើអ្នករកឃើញកំហុស សូមរំលេចអត្ថបទមួយ ហើយចុច បញ្ជា (Ctrl)+បញ្ចូល.